Welcome to IgMin Research – an Open Access journal uniting Biology, Medicine, and Engineering. We’re dedicated to advancing global knowledge and fostering collaboration across scientific fields.
At IgMin Research, we bridge the frontiers of Biology, Medicine, and Engineering to foster interdisciplinary innovation. Our expanded scope now embraces a wide spectrum of scientific disciplines, empowering global researchers to explore, contribute, and collaborate through open access.
Welcome to IgMin, a leading platform dedicated to enhancing knowledge dissemination and professional growth across multiple fields of science, technology, and the humanities. We believe in the power of open access, collaboration, and innovation. Our goal is to provide individuals and organizations with the tools they need to succeed in the global knowledge economy.
IgMin Publications Inc., Suite 102, West Hartford, CT - 06110, USA
Plant polyphenols are ubiquitous secondary metabolites in plants whose antioxidant, anti-inflammatory, antimicrobial, and cardioprotective activities have been systematically elucidated. They exert their physiological functions by scavenging intracellular reactive oxygen species, precisely regulating inflammatory mediators, and inhibiting pathogenic proliferation, exhibiting tremendous application potential in functional foods, biomedicine, and natural cosmetics. However, the inherent chemical instability of polyphenols leads to severe structural degradation and bioactivity loss during extraction, processing, and storage. This manifests not only as detectable content reduction but also as significant "hidden bioactivity loss" without apparent content changes, which has emerged as the core bottleneck restricting their industrial translation. Most existing reviews are limited to single-stage optimizations, and a systematic regulatory framework spanning the entire process of biosynthesis, extraction, and processing has not yet been established. In this narrative critical review, we construct an integrated system for polyphenol content enhancement and stability regulation across the entire value chain from biosynthesis to extraction and processing. We critically elaborate on stress-mediated biosynthetic mechanisms, extraction loss control strategies, and processing stabilization technologies, with the ultimate goal of improving their resource utilization efficiency and advancing the industrial innovation and application of plant polyphenols.
Natural plant polyphenols are a class of secondary metabolites that are ubiquitously distributed and abundant in the plant kingdom, exerting pivotal roles in plant physiological processes, including growth, development, stress resistance, and defense responses. A large number of studies have confirmed that plant polyphenols possess diversified biological activities spanning anti-oxidation [11Divyajanani S, Harithpriya K, Ganesan K, Ramkumar KM. Dietary polyphenols remodel DNA methylation patterns of NRF2 in chronic disease. Nutrients. 2023;15:3347. Available from: https://doi.org/10.3390/nu15153347 ], anti-inflammation [22Qi J, Pan Z, Wang X, Zhang N, He G, Jiang X. Research advances of Zanthoxylum bungeanum polyphenols in inflammatory diseases. Front Immunol. 2024;15:1305886. Available from: https://doi.org/10.3389/fimmu.2024.1305886], anti-bacteria [33Chong Y, Kim BG, Park YJ, Yang Y, Lee SW, Lee Y,et al. Production of four flavonoid C-glucosides in Escherichia coli. J Agric Food Chem. 2023;71:5302-5313. Available from: https://doi.org/10.1021/acs.jafc.3c00297], anti-virus [44Vajdi M, Karimi A, Hassanizadeh S, Farhangi MA, Bagherniya M, Askari G, et al. Effect of polyphenols against complications of COVID-19: current evidence and potential efficacy. Pharmacol Rep. 2024;76:307-327. Available from: https://doi.org/10.1007/s43440-024-00585-6], anti-tumor [55Liu W, Cui X, Zhong Y. Phenolic metabolites as therapeutic in inflammation and neoplasms: molecular pathways explaining their efficacy. Pharmacol Res. 2023;193:106812. Available from: https://doi.org/10.1016/j.phrs.2023.106812], and cardiovascular protection [66Otręba M, Kośmider L, Stojko J, Rzepecka-Stojko A. Cardioprotective activity of selected polyphenols based on epithelial and aortic cell lines: a review. Molecules. 2020;25:5343. Available from: https://doi.org/10.3390/molecules25225343]. These bioactivities are primarily mediated by specific molecular mechanisms: scavenging intracellular reactive oxygen radicals [77Dias MC, Pinto DCGA, Silva AMS. Plant flavonoids: chemical characteristics and biological activity. Molecules. 2021;26:5377. Available from: https://doi.org/10.3390/molecules26175377], precisely regulating inflammatory factor expression [88Medini F, Ksouri R, Msaada K, Legault J. Phenolic compounds from Limonium densiflorum: antioxidant, anti-inflammatory, anticancer, and anti-influenza activities. Int J Environ Health Res. 2025;35:94-104. Available from: https://doi.org/10.1080/09603123.2024.2342572], inhibiting pathogenic microbial proliferation [99Rathod NB, Elabed N, Punia S, Ozogul F, Kim S-K, et al. Recent developments in polyphenol applications on human health: a review. Plants. 2023;12:1217. Available from: https://doi.org/10.3390/plants12061217 ], interfering with tumor cell signaling pathways [1010Moar K, Yadav S, Pant A, et al. Anti-tumor effects of polyphenols via targeting cancer-driving signaling pathways: a review. Indian J Clin Biochem. 2024;39:470-488. Available from: https://doi.org/10.1007/s12291-024-01222-y ], and improving vascular endothelial cell function [1111Serreli G, Deiana M. Role of dietary polyphenols in the activity and expression of nitric oxide synthases: a review. Antioxidants. 2023;12:147. Available from: https://doi.org/10.3390/antiox12010147]. Endowed with these unique physiochemical and biological characteristics, plant polyphenols have become a core research hotspot across food science, pharmacology, and functional materials, and their enormous industrial application potential has garnered extensive global attention. The biosynthesis of plant polyphenols is a sophisticated multi-enzyme synergistic catalytic process dependent on the highly conserved secondary metabolic network in plants, with the shikimic acid and phenylpropanoid pathways as the core [1212Kumar K, Debnath P, Singh S, Kumar N. An overview of plant phenolics and their involvement in abiotic stress tolerance. Stresses. 2023;3:570-585. Available from: https://doi.org/10.3390/stresses3030040]. The phenylpropanoid pathway acts as the key hub governing synthetic efficiency and structural diversity [1313Zhou Z, Duan Y, Li Y, Zhang P, Li Q, Yu L, et al. CYP98A monooxygenases: a key enzyme family in plant phenolic compound biosynthesis. Hortic Res. 2025;12:uhaf074. Available from: https://doi.org/10.1093/hr/uhaf074], which takes L-phenylalanine as the initial precursor and generates p-coumaroyl-CoA via sequential catalysis by rate-limiting enzymes, including phenylalanine ammonia lyase (PAL), cinnamic acid 4-hydroxylase (C4H), and 4-coumarate-CoA ligase (4CL) [1313Zhou Z, Duan Y, Li Y, Zhang P, Li Q, Yu L, et al. CYP98A monooxygenases: a key enzyme family in plant phenolic compound biosynthesis. Hortic Res. 2025;12:uhaf074. Available from: https://doi.org/10.1093/hr/uhaf074]. As the common precursor of flavonoid and non-flavonoid polyphenols, p-coumaroyl-CoA branches into distinct synthetic pathways: flavonoids are formed by enzymatic modification with malonyl-CoA via chalcone synthase (CHS), chalcone isomerase (CHI) and flavanone 3-hydroxylase (F3H), ultimately yielding compounds with the characteristic C6-C3-C6 skeleton [1414Chen S, Wang X, Cheng Y, Gao H, Chen X. A review of classification, biosynthesis, biological activities, and applications of flavonoids. Molecules. 2023;28:4982. Available from: https://doi.org/10.3390/molecules28134982]; non-flavonoids are synthesized through divergent shunts, where p-coumaroyl-CoA undergoes hydroxylation and methylation to form phenolic acids [1515Han S, Cai H, Yu H. UV-C regulation of phenolic biosynthesis in peach fruit during storage. LWT. 2023;190:115573. Available from: https://doi.org/10.1016/j.lwt.2023.115573], polymerization by lignin synthase to produce lignans [1313Zhou Z, Duan Y, Li Y, Zhang P, Li Q, Yu L, et al. CYP98A monooxygenases: a key enzyme family in plant phenolic compound biosynthesis. Hortic Res. 2025;12:uhaf074. Available from: https://doi.org/10.1093/hr/uhaf074], catalysis by stilbene synthase to generate resveratrol and other stilbenes [1616Wang S, Xu Y, Wang F. Postharvest changes in phenolic and volatile compounds in grapes. Food Chem. 2025;465:141958. Available from: https://doi.org/10.1016/j.foodchem.2024.141958], and oligomerization of catechin and epicatechin monomers to form tannins [1717Molnar M, Jakovljević Kovač M, Pavić V. Diversity, structure, biosynthesis, and extraction of tannins using deep eutectic solvents. Molecules. 2024;29:2615. Available from: https://doi.org/10.3390/molecules29112615]. Notably, polyphenol synthesis and accumulation are precisely regulated by both biotic and abiotic stresses: moderate abiotic stresses (light, temperature fluctuation, low salt, heavy metal exposure) modulate the gene transcription and enzyme activity of PAL, CHS and other key synthetases, while biotic stresses (insect herbivory, pathogen infection, weed competition) trigger the jasmonic acid (JA) and salicylic acid (SA) signaling pathways—with ethylene (ET) mediating their crosstalk—to drive defensive polyphenol synthesis. The abscisic acid (ABA) pathway dominates abiotic stress responses, and synergizes with the mitogen-activated protein kinase (MAPK) cascade and reactive oxygen species (ROS) signaling to exert bidirectional regulation on polyphenol biosynthesis and degradation, forming a complex stress regulatory network for polyphenol accumulation.
From a chemical structural perspective, plant polyphenols are defined by at least one aromatic ring with one or more hydroxyl substituents, and are classified into flavonoids and non-flavonoids based on carbon skeleton differences [1818Sharma P, Dhiman T, Negi RS. Molecular mechanisms of skin photoaging and therapeutic advances using polyphenols. S Afr J Bot. 2024;166:466-482. Available from: https://doi.org/10.1016/j.sajb.2024.01.035]. Flavonoids feature a typical C6-C3-C6 skeleton composed of two benzene rings (A and B) linked by a three-carbon bridge, which usually forms a heterocyclic pyran ring; non-flavonoids lack this characteristic skeleton and exhibit greater structural diversity, including phenolic acids (gallic acid, caffeic acid, chlorogenic acid), lignans, stilbenes (resveratrol), and hydrolyzable/condensed tannins (ellagic tannins, proanthocyanidins). Polyphenol chemical stability is predominantly determined by structural characteristics: the number, arrangement, and conjugation degree of phenolic hydroxyl groups. For instance, polyphenols with an o-dihydroxy (catechol) structure (epigallocatechin gallate (EGCG), caffeic acid) are prone to oxidation to form reactive quinones [1919Chrostowski PC, Dietrich AM, Suffet IH. Ozone and oxygen induced oxidative coupling of aqueous phenolics. Water Res. 1983;17:1627-1633. Available from: https://doi.org/10.1016/0043-1354(83)90021-0,2020Spyroudis S. Hydroxyquinones: synthesis and reactivity. Molecules. 2000;5:1291-1330. Available from: https://doi.org/10.3390/51201291]. whereas those with a single phenolic hydroxyl group or sterically hindered hydroxyls exhibit superior stability [2121Alcalde B, Granados M, Saurina J. Exploring the antioxidant features of polyphenols by spectroscopic and electrochemical methods. Antioxidants. 2019;8:523-532. Available from: https://doi.org/10.3390/antiox8110523]. This intrinsic structure-stability relationship creates a core technical paradox for practical application: the same structural features conferring high antioxidant activity render polyphenols highly susceptible to degradation. Throughout their life cycle—biosynthesis, extraction and purification, processing and storage—these degradation pathways cause significant losses in polyphenol content and a sharp decline in bioactivity. More importantly, instability manifests not only as detectable content loss but also as "hidden loss": structural alterations without complete degradation that abrogate bioactivity, further limiting their application value.
Given the scientific and industrial significance of plant polyphenols, a wealth of comprehensive reviews have been published in recent years, each contributing profoundly to advancing our understanding of this complex class of compounds. Numerous authoritative reviews have systematically illustrated the biological activities of polyphenols and their underlying molecular mechanisms, laying a solid foundation for the development of polyphenols as functional food ingredients and pharmaceutical formulations [22–2422–24Bas TG. Dietary polyphenols (flavonoids) derived from plants for therapeutic health: antioxidant performance, ROS, molecular mechanisms, and bioavailability limitations. Int J Mol Sci. 2026;27:1404. Available from: https://doi.org/10.3390/ijms27031404]. Other publications have specifically focused on the biosynthetic pathways and metabolic engineering strategies of polyphenols, delivering theoretical guidance for improving polyphenol yields in planta [25–2725–27Wang Y, Chen J, He G, Yin L, Liao Y. Unlocking the potential of flavonoid biosynthesis through integrated metabolic engineering. Front Plant Sci. 2025;16:1597007. Available from: https://doi.org/10.3389/fpls.2025.1597007]. A further body of research has explored extraction technologies and processing stability, and summarized multiple approaches to alleviate polyphenol degradation during industrial production [28–3028–30Rosa GP, Barreto MC, Seca AML. Ionic liquids and deep eutectic solvents for polyphenol extraction: opportunities and limitations. Int J Mol Sci. 2026;27:3538. Available from: https://doi.org/10.3390/ijms2708353]. Nevertheless, despite these substantial advances, existing reviews are constrained by three major limitations, which create critical knowledge gaps across the field. First, biosynthesis, extraction, processing, and application are commonly regarded as discrete and isolated processes, while the inherent correlations of polyphenol stability challenges throughout the entire value chain are largely unrecognized. Second, most reviews solely address the quantitative losses of polyphenols and largely overlook the aforementioned "latent loss" phenomenon — a more prominent and vastly underappreciated bottleneck in practical applications. Third, the lack of systematic cross-comparison among stability enhancement strategies for different stages hinders researchers and industrial practitioners from screening optimal stage-specific solutions for targeted scenarios.
To resolve the above limitations, this review constructs a full-chain analytical framework for the stability regulation of plant polyphenols, integrating the complete research system covering the full spectrum from biosynthesis to industrial application. Breaking the conventional fragmented research paradigm in this field, the present work elaborates the loss mechanisms and regulatory rules of polyphenols from a full-lifecycle perspective in a unified manner. Unlike previous studies that merely focus on quantitative degradation of polyphenols in a one-sided way, this work takes both apparent losses and latent activity attenuation into consideration, and distills the core mechanisms driving dynamic changes in polyphenol stability. By systematically sorting out the technical deficiencies and industrialization dilemmas in current research, we establish a universal theoretical system for polyphenol stability regulation on the basis of structure-activity relationships.
Regulation Mechanisms of Content in Plant Polyphenol Biosynthesis
Core Biosynthetic Pathways and Key Regulatory Nodes
Dependent on the coordinated action of the shikimate and phenylpropanoid pathways, the biosynthesis of plant polyphenols begins with the shikimate pathway, which transfers carbon flux from primary metabolism to generate aromatic amino acids, including L-phenylalanine and L-tyrosine, through multiple rigorously regulated enzymatic reactions and supplies fundamental precursors for secondary metabolism [3131Tzin V, Malitsky S, Zvi MMB. Expression of bacterial feedback-insensitive DAHP synthase in Arabidopsis and metabolic bottlenecks. New Phytol. 2012;194:430-439. Available from: https://doi.org/:10.1111/j.1469-8137.2012.04052.x,3232Yuan J, Zhong S, Long Y. Shikimate kinase plays important roles in anthocyanin synthesis in Petunia. Int J Mol Sci. 2022;23:15964. Available from: https://doi.org/10.3390/ijms232415964]. Within this pathway, 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase (DAHPS), 3-dehydroquinate synthase (DHQS), and shikimate kinase (SK) function as major regulatory nodes to maintain the homeostasis of precursor supply; as central metabolic switches, these enzymes further control carbon flux allocation to the downstream phenylpropanoid pathway, facilitating polyphenol skeleton assembly, substance conversion and metabolic branching, and ultimately underpinning the entire biosynthetic process of plant polyphenols.
Entry into the phenylpropanoid route is strictly gated by PAL, which commits carbon from L-phenylalanine to the formation of cinnamic acid derivatives; subsequent hydroxylation by C4H and CoA ligation by 4CL converge to generate p-coumaroyl-CoA, the single metabolic hub from which all major polyphenol classes originate [3333Azizyan R, Abdollahi Mandoulakani B. Gene expression and phenolic compounds in Sonchus arvensis under drought stress. Ind Crops Prod. 2024;209:118030. Available from: https://doi.org/10.1016/j.indcrop.2024.118030]. PAL, C4H, and 4CL are recognized as the core rate-limiting enzymes of this pathway, and their catalytic efficiency and expression levels directly determine the yield of p-coumaroyl-CoA, thus regulating the overall biosynthetic efficiency of polyphenols.
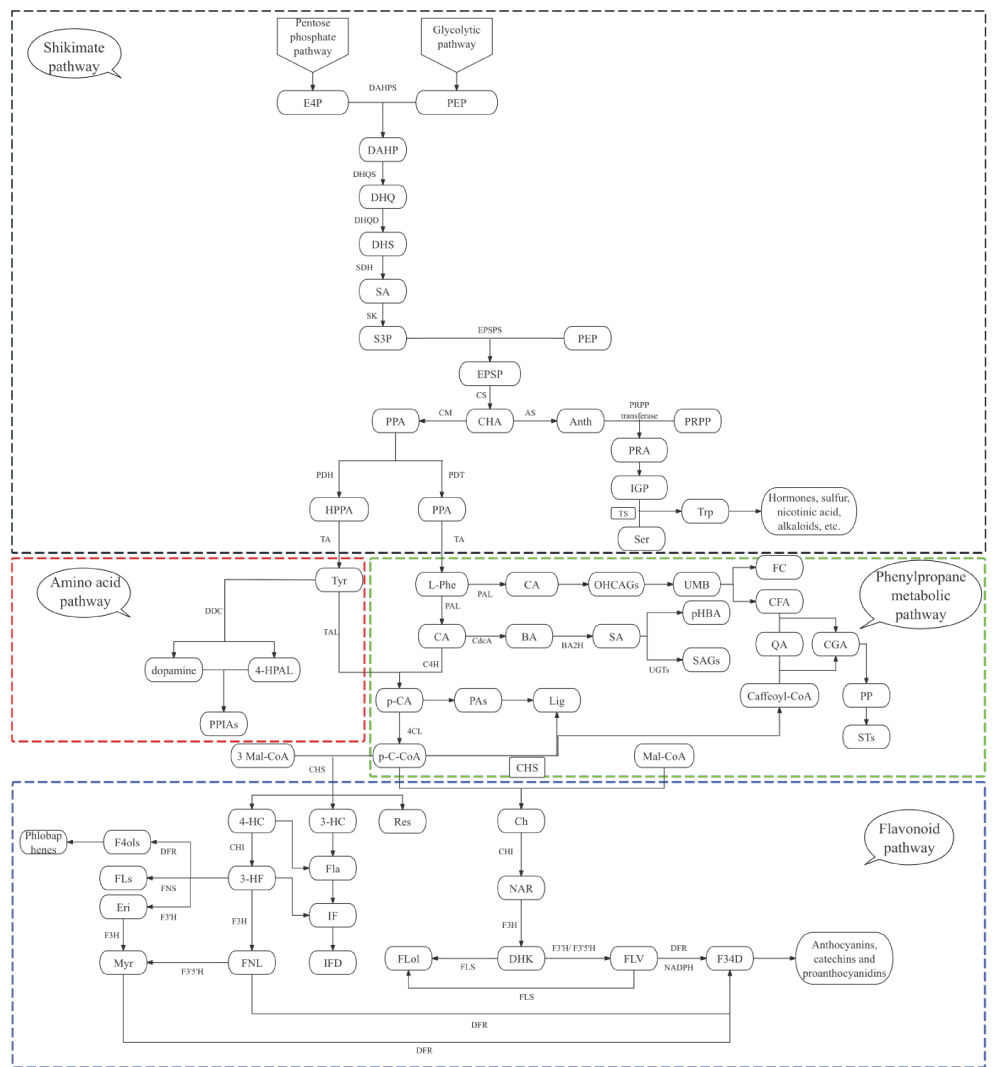
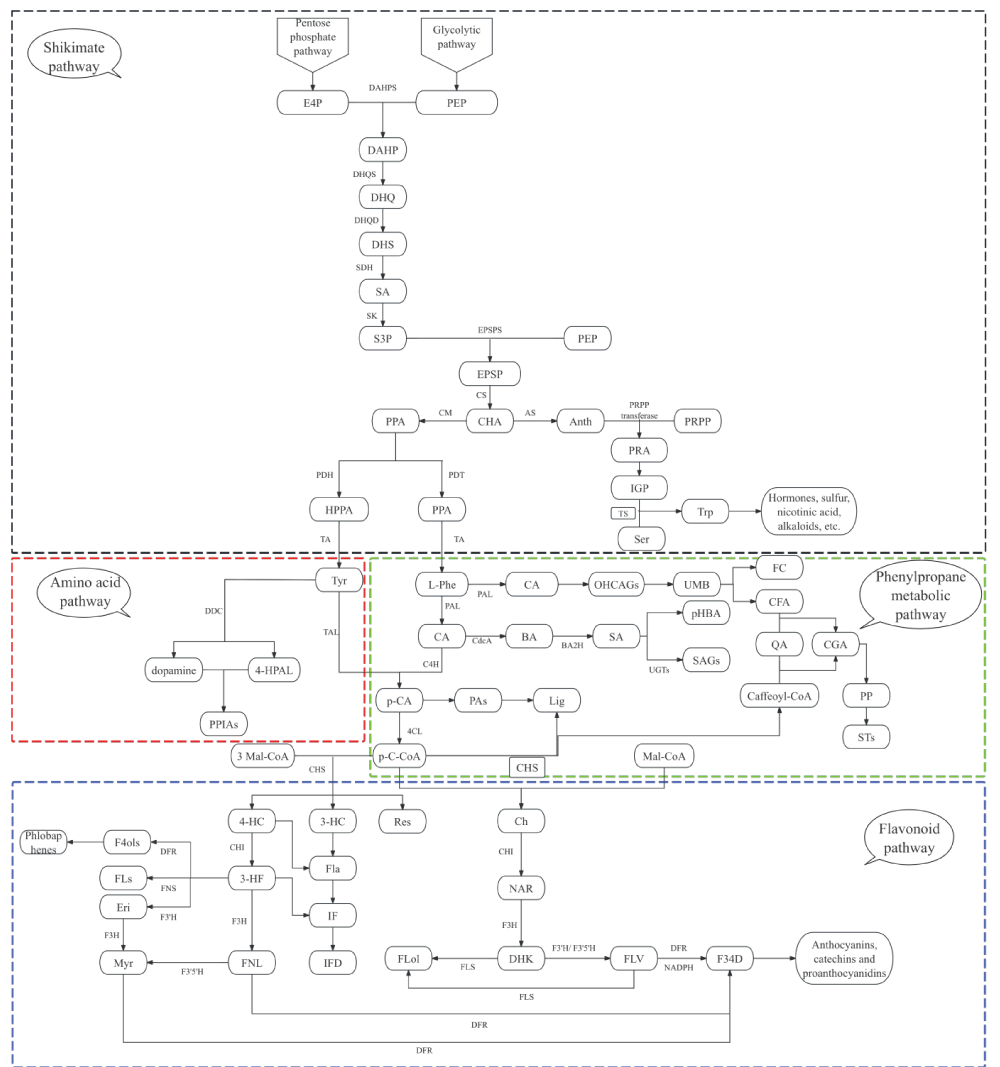
At the central metabolic node of p-coumaroyl-CoA, pathway partitioning is governed by the competitive activities of distinct branch-entry enzymes, which further diverge into flavonoid and non-flavonoid polyphenol biosynthesis. In the flavonoid direction, CHS catalyzes the iterative condensation of p-coumaroyl-CoA with malonyl-CoA to construct the chalcone backbone, while CHI subsequently directs the cyclization toward flavanone intermediates, establishing the C6-C3-C6 scaffold that defines this class [1414Chen S, Wang X, Cheng Y, Gao H, Chen X. A review of classification, biosynthesis, biological activities, and applications of flavonoids. Molecules. 2023;28:4982. Available from: https://doi.org/10.3390/molecules28134982]. Non-flavonoid branches diverge through alternative enzymatic routes: hydroxycinnamoyl-CoA derivatives are channeled into phenolic acid conjugates and lignan biosynthesis via dirigent protein-mediated coupling [3434Yao L, Wu X, Jiang X. Subcellular compartmentalization in biosynthesis and engineering of plant natural products. Biotechnol Adv. 2023;69:108258. Available from: https://doi.org/10.1016/j.biotechadv.2023.10825], stilbene synthase competitively utilizes the same CoA esters to generate resveratrol and its analogs [3535Jeandet P, Sobarzo-Sánchez E, Silva AS, et al. Biocatalytic and green chemistry methods for resveratrol production. Biotechnol Adv. 2020;39:107461. Available from: https://doi.org/10.1016/j.biotechadv.2019.107461], and specific glycosyltransferases or oxidative enzymes orchestrate the assembly of hydrolyzable and condensed tannins [1717Molnar M, Jakovljević Kovač M, Pavić V. Diversity, structure, biosynthesis, and extraction of tannins using deep eutectic solvents. Molecules. 2024;29:2615. Available from: https://doi.org/10.3390/molecules29112615] (Figure 1).
Figure 1: Comprehensive biosynthetic pathway network of plant polyphenols. The diagram illustrates three interconnected modules: (a) the shikimate pathway (upper, black dashed box) generating aromatic amino acid precursors; (b) the core phenylpropanoid pathway (middle, green dashed box) producing p-coumaroyl-CoA; (c) the amino acid pathway (middle, red dashed box); and (d) downstream branch pathways (lower, blue/red boxes) generating diverse polyphenol classes.
Throughout this metabolic network, the magnitude of carbon flux into each terminal polyphenol product is predominantly determined by the expression levels and catalytic activity of branch-specific rate-limiting enzymes. These enzymatic nodes function as both metabolic bottlenecks and regulatory hubs, allowing the system to dynamically adjust polyphenol composition and abundance in response to upstream precursor availability and downstream metabolic demand, thereby establishing the mechanistic framework for understanding how pathway activity governs the quantitative and qualitative output of polyphenol diversity.
Regulatory Mechanisms of Biotic Stress on Polyphenol Biosynthesis
As core defensive secondary metabolites of plants, polyphenol biosynthesis will be dynamically reprogrammed by interconnected hormone signaling and transcriptional regulatory networks once plants encounter biotic stress such as herbivore attack, fungal pathogen infection, and weed competition (Figure 1 and Table 1). The co-evolutionary interplay between plants and biotic stressors further modulates the specificity and response intensity of polyphenol metabolism based on different stress types.
Table 1: Types and main changes of biotic stress and abiotic stress
Under herbivore pressure, exemplified by phloem-feeding insects such as aphids and tea green leafhoppers, plants perceive mechanical damage and herbivore-associated molecular patterns (HAMPs), rapidly triggering the JA and SA signaling cascades [3636Liu Q, Li S, Ding W. Aphid-induced tobacco resistance and salicylic acid changes in rhizosphere. Eur J Plant Pathol. 2020;157:465-483. Available from: https://doi.org/10.1007/s10658-020-02005-w,3737Wang X, Zeng L, Liao Y. Herbivore-induced α-farnesene formation in tea plants. Int J Mol Sci. 2019;20:4151. Available from: https://doi.org/10.3390/ijms20174151]. Typically, chewing and piercing-sucking insects predominantly activate the JA pathway, which upregulates the expression of WRKY and MYB family transcription factors that specifically bind to the promoter regions of polyphenol biosynthetic genes [3838Ji N, Wang J, Li Y. PpWRKY70 involvement in disease resistance via phenylpropanoid pathway in peach. Postharvest Biol Technol. 2021;174:111466. Available from: https://doi.org/10.1016/j.postharvbio.2021.111466,3939Xie M, Zhang J, Yao T. ANGUSTIFOLIA regulates MYB46 and WRKY33 transcription in Arabidopsis. New Phytol. 2020;228:1627-1639. Available from: https://doi.org/:10.1111/nph.16826]. Consequently, the activities of core rate-limiting enzymes—particularly PAL and CHS—increase markedly within hours to days post-infestation, driving the accumulation of defensive polyphenols including flavonoids, chlorogenic acid, and condensed tannins [3838Ji N, Wang J, Li Y. PpWRKY70 involvement in disease resistance via phenylpropanoid pathway in peach. Postharvest Biol Technol. 2021;174:111466. Available from: https://doi.org/10.1016/j.postharvbio.2021.111466,4040Hodaei M, Rahimmalek M, Arzani A. Water stress effects on flavonoid biosynthesis in chrysanthemum. Ind Crops Prod. 2018;120:295-304. Available from: https://doi.org/10.1016/j.indcrop.2018.04.07,4141Ding X, Zhu X, Zheng W. BTH treatment delays senescence of postharvest pitaya fruit via antioxidant system and phenylpropanoid pathway. Foods. 2021;10:846. Available from: https://doi.org/10.3390/foods10040846]. This induced resistance not only deters further insect feeding but also reduces pest fecundity, representing a classic example of herbivore-triggered metabolic reinforcement in the phenylpropanoid pathway [4242Mouden S, Bac-Molenaar JA, Kappers IF. Elicitor application in strawberry increases plant resilience without yield loss. Front Plant Sci. 2021;12:695908. Available from: https://doi.org/10.3389/fpls.2021.695908].
Fungal pathogens, including soil-borne Rhizoctonia solani and Fusarium oxysporum, elicit distinct yet overlapping defense responses compared with herbivore stress [4343Manoharan B, Qi SS, Dhandapani V. Gene expression profiling reveals enhanced defense responses in invasive weed vs native congener. Int J Mol Sci. 2019;20:4916. Available from: https://doi.org/10.3390/ijms20194916,4444Duhan L, Kumar D, Pasrija R. Exogenous salicylic acid mitigates Fusarium stress in Vigna mungo. Plant Cell Rep. 2025;44:2. Available from: https://doi.org/10.1007/s00299-024-03394-6]. Upon fungal pathogen-associated molecular patterns (PAMPs) recognition, plants initiate a biphasic SA-JA signaling dynamic: an early SA burst that promotes lignification and the accumulation of hydroxycinnamic acids at infection sites to block pathogen invasion, followed by a JA-mediated response that reinforces systemic acquired resistance (SAR) [4545Guo M, Hou J, Li C. Calcium signaling mediates lignin synthesis in pear exocarp. Plant Physiol Biochem. 2022;190:174-183. Available from: https://doi.org/10.1016/j.plaphy.2022.09.00 ]. In this process, polyphenol biosynthesis is redirected toward the production of lignin monomers, phenolic acids, and condensed tannins, which not only strengthen plant cell walls as a physical barrier but also exert direct antifungal activity [4646Zeng Y, Song H, Xia L. Defensive responses of poplars to fungal pathogens. Front Plant Sci. 2023;14:1107583. Available from: https://doi.org/10.3389/fpls.2023.1107583]. The sustained upregulation of PAL, C4H, and 4CL during the entire infection period ensures a continuous supply of phenylpropanoid precursors for both structural reinforcement and chemical inhibition of pathogens [4747Pant SR, Irigoyen S, Liu J. Phenylalanine ammonia lyase promotes antiviral defenses in Brachypodium. mBio. 2021;12:e03518-20. Available from: https://doi.org/10.1128/mBio.03518-20]. Notably, the ET signaling pathway mediates the crosstalk between SA and JA pathways here, fine-tuning the dynamic balance between local and systemic defense responses to avoid excessive metabolic consumption [3939Xie M, Zhang J, Yao T. ANGUSTIFOLIA regulates MYB46 and WRKY33 transcription in Arabidopsis. New Phytol. 2020;228:1627-1639. Available from: https://doi.org/:10.1111/nph.16826].
Weed competition imposes a more complex stress regime involving both resource deprivation and allelopathic interference. In crops such as peach and buckwheat, competition for light, nutrients, and water induces endogenous oxidative stress, while allelochemicals released by neighboring weeds further amplify ROS production in plant tissues [4848Carbonaro M, Mattera M. Polyphenoloxidase activity in organic vs conventional fruits. Food Chem. 2001;72:419-424. Available from: https://doi.org/10.1016/S0308-8146(00)00248-X,4949Merino J, Pedreros A, Fischer S. Weed interference affects polyphenol content in quinoa. Chil J Agric Res. 2019;79:405-414. Available from: https://doi.org/10.4067/S0718-58392019000300405]. This sustained redox imbalance activates the phenylpropanoid metabolic pathway as an antioxidant countermeasure, resulting in the upregulation of CHS and downstream branch enzymes, as well as elevated levels of phenolic acids, flavonoids, and proanthocyanidins [5050Tyagi K, Maoz I, Kochanek B. Cytokinin effects on phenylpropanoid pathway in grape. Hortic Res. 2021;8:51-65. Available from: https://doi.org/10.1038/s41438-021-00488-0,5151Chen Z, Zhang L, Peng M. Selenite enhances Chinese flowering cabbage quality via antioxidant regulation. Food Res Int. 2023;163:112229. Available from: https://doi.org/10.1016/j.foodres.2022.112229]. The metabolic reprogramming under weed competition highlights the dual role of polyphenols: on the one hand, they scavenge excess ROS to mitigate oxidative damage, and on the other hand, they mediate plant-plant allelopathic interactions to adapt to the competitive environment [5252Reshi ZA, Ahmad W, Lukatkin AS. Secondary metabolite biosynthesis pathways and environmental influences. Metabolites. 2023;13:895. Available from: https://doi.org/10.3390/metabo13080895,5353Yeshi K, Crayn D, Ritmejerytė E. Secondary metabolites under abiotic stress and pharmaceutical applications. Molecules. 2022;27:313. Available from: https://doi.org/10.3390/molecules27010313].
The regulatory specificity of biotic stress on polyphenol biosynthesis lies in the non-linear relationship between stress type, signaling pathway, and polyphenol metabolic output. Different biotic attackers trigger unique transcriptional signatures in plants, leading to the accumulation of distinct polyphenol profiles: JA-dominant herbivore responses favor the synthesis of flavonoids and condensed tannins, whereas SA-dominant fungal infection responses prioritize the production of lignin monomers and simple phenolic acids. Moreover, the same stressor can elicit divergent biosynthetic responses across different plant species and genotypes, reflecting evolutionary adaptations in receptor specificity and metabolic pathway wiring.
Collectively, this section systematically elucidates the mechanisms underlying the dynamic reprogramming of polyphenol biosynthesis in plants exposed to three typical biotic stresses. It reveals the specific crosstalk patterns of jasmonic acid (JA), salicylic acid (SA), and ethylene (ET) signaling pathways, along with their core roles in shaping stress-specific polyphenol metabolic characteristics. The stress type–signaling pathway–polyphenol response regulatory axis constructed in this study clarifies the core mechanisms of plant adaptive chemical defenses against various biotic stresses. These findings not only advance the understanding of plant metabolic adaptation and the co-evolutionary mechanisms between plants and biotic stressors, but also provide precise and practically applicable molecular targets for artificial regulation of the phenylpropanoid pathway and improvement of stress tolerance in crops.
Regulatory Mechanisms of Abiotic Stress on Polyphenol Biosynthesis
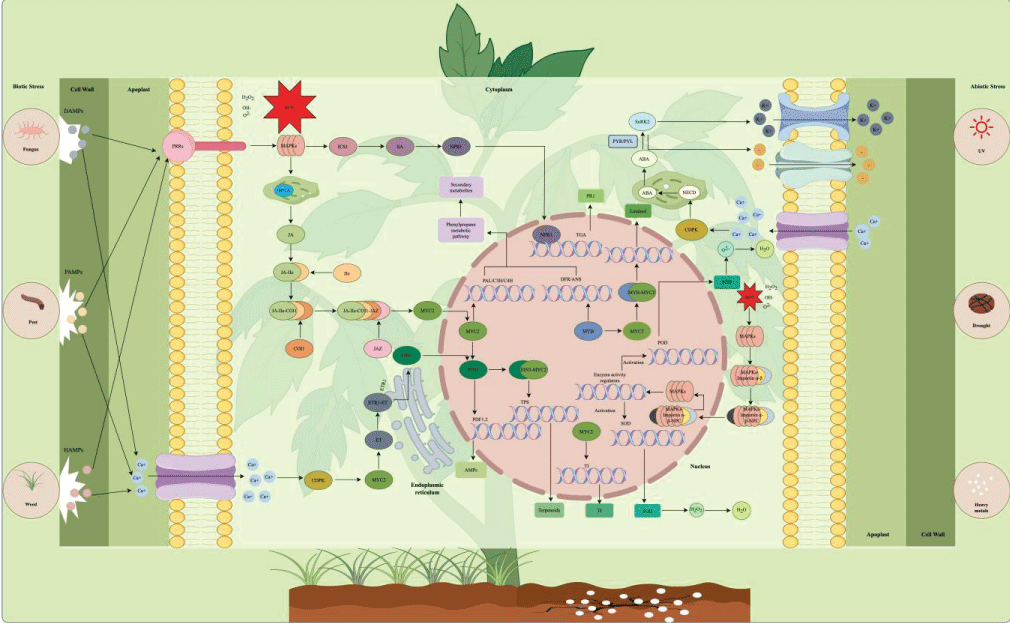
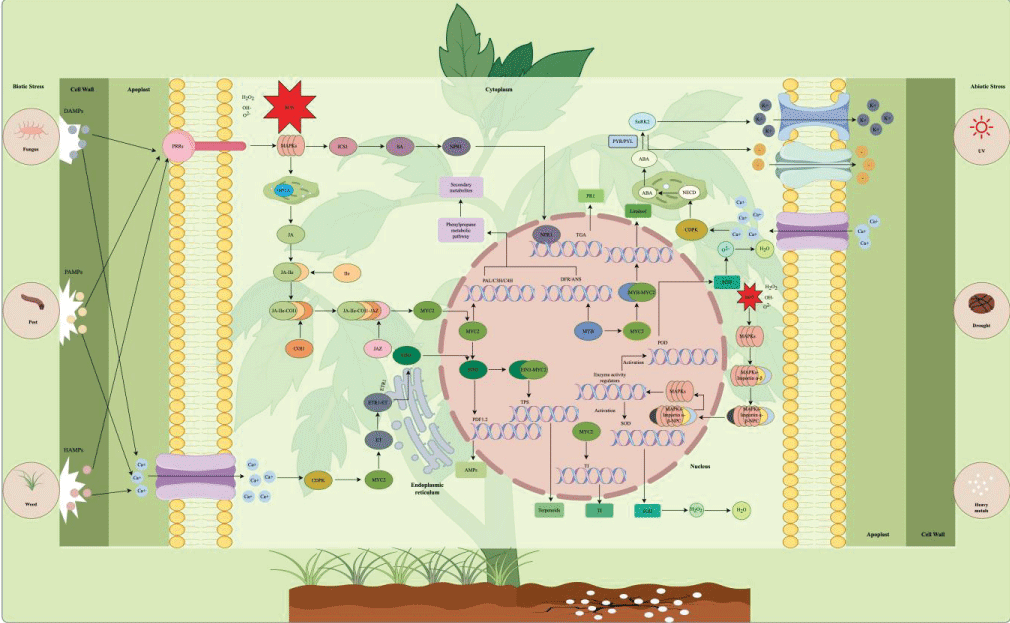
Abiotic stresses, including drought, salinity, ultraviolet (UV) radiation, heavy metal toxicity, and thermal stress, are major environmental drivers that reshape plant secondary metabolic flux. Unlike biotic stresses, which trigger pathogen-specific defense responses primarily mediated by JA and SA signaling (detailed in Section 1.2), abiotic stresses elicit global adaptive metabolic adjustments centered on ABA signaling, making it a well-established, controllable strategy for artificially enhancing the endogenous polyphenol pool in plant raw materials via targeted pre-harvest elicitation (Figure 2 and Table 1).
Figure 2: Signaling regulatory network of polyphenol biosynthesis under biotic and abiotic stresses. Left panel: biotic stress perception (herbivory, pathogens, weeds) triggers JA/SA/ET signaling cascades that activate transcription factors (WRKY, MYB) binding to biosynthetic gene promoters. Right panel: abiotic stresses (drought, salinity, UV, heavy metals) converge on ABA-mediated signaling, integrating MAPK cascades and ROS signaling to modulate polyphenol accumulation.
Single-factor abiotic stress treatments exert consistent biphasic, dose-dependent effects on polyphenol accumulation across a wide range of edible and medicinal plant species. Under drought conditions, mild to moderate water deficit typically upregulates polyphenol biosynthesis: it activates ABA-dependent signaling cascades, induces the transcriptional expression of core rate-limiting enzymes (PAL and CHS), and enhances the flux of the shikimate and phenylpropanoid pathways, leading to significant accumulation of flavonoids, phenolic acids, and condensed tannins in tea plant and sesametea [5454Wang Y, Fan K, Wang J. Proteomic analysis of Camellia sinensis under drought stress. J Plant Physiol. 2017;219:91-99. Available from: https://doi.org/10.1016/j.jplph.2017.10.001,5555Ghotbzadeh Kermani S, Saeidi G, Sabzalian MR. Drought stress influences sesamin and polyphenols in sesame. Food Chem. 2019;289:360-368. Available from: https://doi.org/10.1016/j.foodchem.2019.03.004]. In contrast, severe and prolonged drought suppresses photosynthetic carbon assimilation and primary metabolic activity, limiting the precursor supply for polyphenol synthesis and ultimately reducing total polyphenol yield. Salinity stress follows an identical biphasic pattern: low to moderate salt concentrations stimulate the accumulation of flavonoids and proanthocyanidins, which act as compatible osmoprotectants and ROS scavengers, while high salinity causes irreversible cellular damage and inhibits secondary metabolism [5656Xu N, Liu S, Lu Z. Flavonoid accumulation under salt stress in Ginkgo biloba. Plants. 2020;9:1162. Available from: https://doi.org/10.3390/plants9091162]. UV-B radiation is the most potent abiotic elicitor for flavonoid biosynthesis, particularly for anthocyanins and flavonols, which function as UV-absorbing sunscreens and antioxidants to mitigate photodamage; this effect has been widely validated in leafy vegetables and timber trees [57–5957–59Fraser DP, Sharma A, Fletcher T. UV-B increases flavonoid quercetin in coriander. Sci Rep. 2017;7:17758. Available from: https://doi.org/10.1038/s41598-017-18073-8]. Heavy metal stress (e.g., cadmium, excess copper) and thermal stress also trigger polyphenol accumulation, primarily through the activation of MAPK cascades and ROS burst, with ABA serving as the central hormonal hub that integrates multiple stress signals and coordinates the transcriptional activation of core polyphenol biosynthetic genes [60–6360–63Izbiańska K, Arasimowicz-Jelonek M, Deckert J. Phenylpropanoid metabolites improve lupine root tolerance to lead stress. Ecotoxicol Environ Saf. 2014;110:61-67. Available from: https://doi.org/:10.1016/j.ecoenv.2014.08.014].
A defining feature of abiotic stress regulation is its bidirectional, intensity-dependent effect on polyphenol metabolism, which is universally described by the "eustress-distress" framework in plant stress physiology. Mild to moderate stress (eustress) induces adaptive, non-damaging responses: transient ROS elevation acts as a secondary messenger to activate antioxidant polyphenol synthesis, and moderate ABA accumulation upregulates phenylpropanoid pathway genes without compromising primary metabolism and plant growth. Conversely, severe or prolonged stress (distress) causes metabolic collapse, characterized by irreversible oxidative damage, reduced photosynthetic capacity, and downregulation of biosynthetic genes, leading to simultaneous losses in plant biomass and polyphenol content. This duality establishes a clear operational window for pre-harvest elicitation: only stress intensities within the eustress range can achieve the dual goal of maintaining plant yield and enhancing polyphenol accumulation. The temporal dimension of stress regulation is equally critical: short-term stress exposure often elicits rapid and reversible transcriptional activation of biosynthetic genes, while chronic stress leads to negative feedback inhibition and metabolic exhaustion, which is often overlooked in current laboratory studies.
Despite these limitations, this network provides a core theoretical blueprint for rational source-based polyphenol enhancement, laying a high-quality feedstock foundation for downstream processing. Notably, substantial polyphenol loss during extraction remains the next core bottleneck for industrial translation, with underlying mechanisms and mitigation strategies systematically addressed in Section 2.
Loss Mechanisms During Polyphenol Extraction and Strategies for Quality Improvement and Loss Reduction
Extraction serves as the pivotal hub linking in planta polyphenol biosynthesis to their end-point high-value valorization. The selection of extraction technical route directly determines the yield, purity, structural integrity, and bioactivity retention of polyphenols released from plant matrices, and further dominates their subsequent processing stability, oral bioavailability, and industrial translation potential. Currently, extraction techniques for plant polyphenols are broadly categorized into two major groups: conventional extraction methods and novel intensified extraction methods [7272Sridhar A, Ponnuchamy M, Kumar PS. Techniques and modeling of polyphenol extraction: a review. Environ Chem Lett. 2021;19:3409-3443. Available from: https://doi.org/10.1007/s10311-021-01217-8] (Table 2). However, all existing extraction techniques are accompanied by inevitable degradation and activity loss of polyphenols, whose underlying mechanisms and targeted mitigation strategies are systematically dissected in this chapter.
Table 2: Advanced extraction technology and the yield of polyphenols from different sources
Influencing Factors of Polyphenol Extraction
As the critical bridge between in planta polyphenol biosynthesis and downstream high-value utilization, extraction directly determines the final yield and functional quality of polyphenol products. Polyphenol loss during this process arises from irreversible structural disruption and concomitant bioactivity attenuation, which are mainly mediated by three core chemical pathways: oxidative degradation, hydrolysis, and isomerization.
Oxidative degradation represents the predominant cause of polyphenol loss during extraction, and the degree of such degradation varies markedly among different extraction methods. For plant materials including grapes, vanilla, cabbages and cereals, conventional solvent extraction performed under atmospheric pressure, open systems, prolonged extraction duration, continuous exposure to oxygen and light, and without deoxygenation or light shielding leads to 41%–59% thermal oxidation loss of flavonoids and anthocyanins within polyphenol fractions [8080Antony A, Farid M. Effect of temperature on polyphenol extraction. Appl Sci. 2022;12:2107. Available from: https://doi.org/10.3390/app12042107]. When flesh tissues of Fuji apple are used as raw materials, high-shear homogenization with a traditional high-speed blender severely disrupts cellular structures and triggers massive release of polyphenol oxidase. The subsequent aerobic environment induces intense enzymatic oxidation, resulting in a total polyphenol loss of 60.0% and total flavonoid degradation of 66.9% [8181Park SY, Kang TM, Kim MJ. Enzymatic browning reaction of apple juices prepared using different juicing systems. Biosci Biotechnol Biochem. 2018;82:2000-2006. Available from: https://doi.org/10.1080/09168451.2018.149794]. For mixed plant matrices composed of chamomile, artichoke and coffee beans, oxidation readily occurs during conventional microwave-assisted and ultrasound-assisted extraction. In these cases, the maximum loss of monocaffeoylquinic acid reaches approximately 30%, while the degradation rate of dicaffeoylquinic acid can be as high as 60% [8282Olszowy-Tomczyk M, Paprotny Ł, Wianowska D. Stability of phenolic acids under extraction conditions. Molecules. 2024;29:5861. Available from: https://doi.org/10.3390/molecules29245861]. The molecular mechanism involves three sequential steps: (1) chelation of Fe3+/Cu2+ with the ortho-dihydroxy group forming a five-membered ring complex; (2) electron transfer from polyphenol to metal ion generating semiquinone radicals; (3) radical chain propagation leading to quinone formation and subsequent polymerization. Conventional methods, such as heating reflux and Soxhlet extraction, synergistically accelerate this process through prolonged heating, oxygen exposure, and transition metal catalysis [8383Wang G, Kumar Y. Mechanisms of initial non-enzymatic oxidation of wine: a mini review. J Food Sci. 2024;89:2530-2545. Available from: https://doi.org/10.1111/1750-3841.17038]. Meanwhile, these methods also trigger lipid oxidation via the Fenton reaction, resulting in nutrient loss and deterioration of food quality [8484Mussio C, Garcia-Perez P, Moret E. Natural chelating agents in food stability: mechanisms and applications. Food Chem. 2025;496:146682. Available from: https://doi.org/10.1016/j.foodchem.2025.14668 .]. While novel extraction technologies significantly improve extraction efficiency, they introduce new oxidative degradation risks: ultrasound-assisted extraction (UAE) generates highly reactive hydroxyl radicals via cavitation-induced water sonolysis [8585Wang P, Cheng C, Ma Y. Degradation of polyphenols in aqueous extraction systems under ultrasound. Sep Purif Technol. 2020;247:116967. Available from: https://doi.org/10.1016/j.seppur.2020.116967]; microwave-assisted extraction (MAE) promotes overheating-driven free radical reactions; enzyme-assisted extraction (EAE) carries polyphenol oxidase (PPO)-catalyzed enzymatic oxidation and browning risk [8686Arnold M, Gramza-Michałowska A. Enzymatic browning in apple products: inhibition strategies. Compr Rev Food Sci Food Saf. 2022;21:5038-5076. Available from: https://doi.org/10.1111/1541-4337.13059,8787Tilley A, McHenry MP, McHenry JA. Role of substrates in polyphenol oxidase-mediated browning. Curr Res Food Sci. 2023;7:100623. Available from: https://doi.org/10.1016/j.crfs.2023.100623].
Hydrolysis is a ubiquitous core reaction during polyphenol extraction, directly governing the structural composition and bioactivity of polyphenol products. Over 80% of polyphenols in most edible and medicinal plant matrices exist in glycosylated forms [8888Ebrahimi P, Bayram I, Lante A. Phenolic extract of parsley inhibits lipid oxidation in emulsions. Food Res Int. 2024;187:114452. Available from: doi:10.1016/j.foodres.2024.114452.]. Phenolic aglycones, covalently linked to sugar moieties via O-glycosidic bonds, exhibit high intracellular bioactivity but poor water solubility and chemical stability, making them highly susceptible to secondary degradation [8888Ebrahimi P, Bayram I, Lante A. Phenolic extract of parsley inhibits lipid oxidation in emulsions. Food Res Int. 2024;187:114452. Available from: doi:10.1016/j.foodres.2024.114452.]. In conventional thermal reflux and Soxhlet extraction, high temperature cleaves O-glycosidic bonds due to their thermodynamic instability and modulates hydrolysis by altering solvent physicochemical properties [88–9088–90Ebrahimi P, Bayram I, Lante A. Phenolic extract of parsley inhibits lipid oxidation in emulsions. Food Res Int. 2024;187:114452. Available from: doi:10.1016/j.foodres.2024.114452.]; strong acidic or alkaline conditions mediate O-glycosidic bond cleavage and ester hydrolysis respectively via nucleophilic attack [8888Ebrahimi P, Bayram I, Lante A. Phenolic extract of parsley inhibits lipid oxidation in emulsions. Food Res Int. 2024;187:114452. Available from: doi:10.1016/j.foodres.2024.114452.,9191Ikeda T, Masuda T, Takayama M. Solvent-induced emission of organogels. Org Biomol Chem. 2016;14:36-39. Available from: https://doi.org/10.1039/C5OB01898F]. UAE and MAE promote glycosidic bond cleavage via localized high-temperature effects [9292Herrera-Pool E, Ramos-Díaz AL, Lizardi-Jiménez MA. Solvent polarity in ultrasound-assisted extraction of phenolics from habanero pepper leaves. Ultrason Sonochem. 2021;76:105658. Available from: https://doi.org/10.1016/j.ultsonch.2021.105658,9393Wibisono Y, Rachmawati SA, Mylani VS. Cellulose acetate membrane with Olea europaea nanosolids. Alexandria Eng J. 2023;64:119-129. Available from: https://doi.org/10.1016/j.aej.2022.08.036]; EAE precisely hydrolyzes O-glycosidic bonds to release free aglycones through glycosidase-specific catalysis [9494Dilokpimol A, Mäkelä MR, Aguilar-Pontes MV. Diversity of fungal feruloyl esterases and applications. Biotechnol Biofuels. 2016;9:231. Available from: https://doi.org/10.1186/s13068-016-0651-6,9595Gulsunoglu-Konuskan Z, Kilic-Akyilmaz M. Microbial bioconversion of phenolic compounds in agro-industrial wastes. J Agric Food Chem. 2022;70:6901-6910. Available from: https://doi.org/10.1021/acs.jafc.1c06888].
Elevated temperature and light induce polyphenol conformational changes and isomerization, exemplified by the conversion of bioactive trans-resveratrol to low-activity cis-isomer, which impairs biological functionality. Extreme pH alters polyphenol ionization, electron cloud distribution and bond stability, accelerating isomerization and degradation [9696Méndez DA, Fabra MJ, Odriozola-Serrano I. Extraction effects on pectin and phenolics from persimmon waste streams. Food Hydrocolloids. 2022;123:107066. Available from: https://doi.org/10.1016/j.foodhyd.2021.107066]; for instance, anthocyanins are stable under acidic conditions but undergo structural rearrangement and discoloration in neutral/alkaline environments [9797Mahamoud R, Bowman DT, Ward WE. Stability of polyphenols in red rooibos tea. Food Chem. 2024;448:139068. Available from: https://doi.org/10.1016/j.foodchem.2024.139068,9898Fu X, Du Y, Zou L, et al. Acidified glycerol extraction and stabilization of blueberry anthocyanins. Food Chem. 2022;390:133226. Available from: https://doi.org/10.1016/j.foodchem.2022.133226]. Additionally, metal ions catalyze polyphenol oxidation and isomerization, and form chelates with phenolic hydroxyl groups to promote conformational alterations [9999Hosseini Taheri SE, Bazargan M, Rahnama Vosough P. Composition and oxidation of peanut: a review. J Food Compos Anal. 2024;125:105770. Available from: https://doi.org/10.1016/j.jfca.2023.105770]. Common extraction technologies also induce structural alterations: UAE generates reactive oxygen species for isomerization [100100Alean J, Chejne F, Rojano B. Degradation of polyphenols during cocoa drying. J Food Eng. 2016;189:99-105. Available from: https://doi.org/10.1016/j.jfoodeng.2016.05.026], while EAE’s glycoside hydrolases may indirectly cause free polyphenol isomerization/degradation [101101Leangnim N, Unban K, Thangsunan P. Ultrasonic-assisted enzymatic improvement of polyphenol content and antioxidant activity of miang extracts. Ultrason Sonochem. 2023;94:106351. Available from: https://doi.org/10.1016/j.ultsonch.2023.106351].
Key extraction process parameters directly modulate the occurrence and intensity of the above degradation pathways. Prolonged high-temperature treatment, a core limitation of conventional extraction methods, synergistically accelerates all three degradation reactions, reducing both polyphenol recovery yield and antioxidant activity retention. Oxygen exposure drives oxidative polymerization, while inappropriate pH promotes hydrolysis and isomerization . High-concentration organic solvents, despite improving mass transfer, may induce solvent-mediated structural destabilization of sensitive phenolics. These cumulative losses ultimately attenuate the health-promoting bioactivities of polyphenol products, limiting their downstream application. Elucidating these loss mechanisms provides a critical theoretical basis for developing targeted mitigation strategies to maximize polyphenol extraction efficiency and bioactivity retention.
Loss Reduction Strategies for Polyphenol Extraction Processes
At the process parameter level, a low-temperature and short-time extraction strategy effectively inhibits the degradation of heat-sensitive polyphenols by reducing thermal isomerization and hydrolysis, while maintaining acceptable extraction efficiency [102102Chen J, Wang H. Processing characteristics of chia seed: changes in physicochemical properties and structure. Food Biosci. 2024;59:103813. Available from: https://doi.org/10.1016/j.fbio.2024.103813]. For the solvent system, ethanol remains the most widely used solvent for plant polyphenols, providing satisfactory performance with balanced polarity and safety [103103Kotsou K, Stoikou M, Athanasiadis V. Optimization of extraction to enhance antioxidant properties of Prunus spinosa Horticulturae. 2023;9:942-957.Available from: https://doi.org/10.3390/horticulturae9080942,104104Chamali S, Bendaoud H, Bouajila J. Optimization of accelerated solvent extraction of bioactive compounds from Eucalyptus intertexta. J Appl Res Med Aromat Plants. 2023;35:100464. Available from: https://doi.org/10.1016/j.jarmap.2023.100464]. Meanwhile, controlling the system pH within a weakly acidic range (pH 4–6) helps preserve the structural stability of polyphenol glycosides and alleviate base-catalyzed hydrolysis and isomerization [105105Wang D, Wang Y, Zhang Z. Degradation and isomerization of dicaffeoylquinic acids under ultrasound. Ultrason Sonochem. 2023;95:106401. Available from: https://doi.org/10.1016/j.ultsonch.2023.106401,106106Xie C, Yu K, Zhong D, Yuan T, Ye F, Jarrell JA, et al. Isomeric transformations of chlorogenic acid under different conditions. J Agric Food Chem. 2011;59:11078-11087. Available from: https://doi.org/10.1021/jf203104k]. To mitigate oxidation—the primary pathway for polyphenol loss during extraction—inert gas (e.g., nitrogen) protection significantly reduces dissolved oxygen and inhibits oxidative polymerization [107107Gao Y, Song O, Wang M, Guo X, Zhang G, Liu X, et al. Hydrogen protection enhances bioactivity of chrysanthemum extract. Antioxidants. 2023;12:1019. Available from: https://doi.org/10.3390/antiox12051019,108108Wagner A, Dussling S, Scansani S, Bach P, Ludwig M, Steingass CB, et al. Juice production systems and storage effects in red-fleshed apples. Molecules. 2022;27:2459. Available from: https://doi.org/10.3390/molecules27082459]. The addition of suitable antioxidants (e.g., vitamin C, citric acid) further scavenges free radicals, blocks oxidative chain reactions, and protects the phenolic hydroxyl groups from damage [109109Brewer MS. Natural antioxidants: sources, mechanisms, and applications. Compr Rev Food Sci Food Saf. 2011;10:221-247. Available from: https://doi.org/10.1111/j.1541-4337.2011.00156,110110Losada-Barreiro S, Sezgin-Bayindir Z, Paiva-Martins F, Bravo-Díaz C. Biochemistry of antioxidants: mechanisms and applications. Biomedicines. 2022;10:3051. Available from: https://doi.org/10.3390/biomedicines10123051]. These strategies are easy to implement without major equipment modification and show excellent industrial compatibility.
Application and Effects on Quality Improvement and Loss Reduction of Novel Intensified Extraction Technologies
Novel intensified extraction technologies enhance polyphenol recovery through physical field intensification or biocatalysis, while effectively suppressing activity loss. Representative techniques include UAE, MAE, high-pressure-assisted extraction (HPAE), pulsed electric field extraction (PEF), EAE, supercritical fluid extraction (SFE), and natural deep eutectic solvent (NADES)-based extraction.
Physical field-enhanced methods (UAE, MAE, HPAE, PEF) disrupt plant tissue structures via external energy input, strengthening mass transfer under mild conditions. UAE relies on cavitation-induced mechanical rupture to release polyphenols rapidly at low temperatures, thus markedly reducing thermal degradation. MAE achieves rapid volumetric heating through dielectric effects, drastically shortening extraction duration and limiting oxidation of heat-sensitive polyphenols. HPAE and PEF permeabilize cell membranes using high pressure or short electric pulses, promoting solvent diffusion and boosting efficiency at
room or mildly elevated temperatures. Biocatalytic strategies, especially EAE, specifically degrade cell wall polysaccharides and disrupt polyphenol-macromolecule bindings under gentle conditions. This allows efficient release of bound polyphenols while maximally preserving their native structure and bioactivity. Fermentation-assisted extraction further integrates matrix decomposition, polyphenol liberation, and biotransformation, leading to improved bioaccessibility and functionality. Green solvent systems represented by SFE and NADES offer sustainable and safe alternatives. SFE using supercritical CO2 realizes solvent-free extraction, although polar modifiers are often needed for hydrophilic polyphenols [111111Vladić J, Jakovljević Kovač M, Pavić V. Green biomass valorization using supercritical fluids and deep eutectic solvents. Antibiotics. 2023;12:1031. Available from: https://doi.org/10.3390/antibiotics12061031,112112Lojková L, Pluháčková H, Benešová K. Trends in quercetin extraction methods. TrAC Trends Anal Chem. 2023;167:117229. Available from: https://doi.org/10.1016/j.trac.2023.11722]. NADES provide a low-toxicity, biodegradable medium with tunable polarity, making them suitable for various classes of polyphenols [113113García-Roldán A, Piriou L, Jauregi P. Natural deep eutectic solvents for polyphenol extraction from coffee grounds. Front Plant Sci. 2023;13:1072592. Available from: https://doi.org/10.3389/fpls.2022.1072592].
Comparative studies have confirmed the significant advantages of these technologies over conventional methods. The UAE extraction yield of olive polyphenols reached 92.49 mg/g, with good retention of antioxidant activity [7373Dobrinčić A, Repajić M, Garofulić IE. Comparison of extraction methods for olive leaf polyphenols. Processes. 2020;8:1008. Available from: https://doi.org/10.3390/pr8091008]. The PEF-ultrasound coupled extraction of litchi pericarp polyphenols achieved an extraction yield of 109.21 mg/g, which was 2.30 times higher than that of conventional hot water extraction [7676Tan MJ, Li Y, Zhao SQ. Synergistic ultrasound and pulsed electric field extraction of litchi peel polyphenols. Int J Biol Macromol. 2024;260:129613. Available from: https://doi.org/10.1016/j.ijbiomac.2024.129613]. In one study, high-pressure-assisted extraction of phenolic compounds from dates yielded a total phenolic content of 257.93 ± 2.77 mg GAE/100g [5151Chen Z, Zhang L, Peng M. Selenite enhances Chinese flowering cabbage quality via antioxidant regulation. Food Res Int. 2023;163:112229. Available from: https://doi.org/10.1016/j.foodres.2022.112229]. In another study, SFE of polyphenols from chili peppers achieved an extraction yield of 1870 mg GAE/100 g extract under conditions of 63.1℃, 1161.82 psi, and 132 min [5353Yeshi K, Crayn D, Ritmejerytė E. Secondary metabolites under abiotic stress and pharmaceutical applications. Molecules. 2022;27:313. Available from: https://doi.org/10.3390/molecules27010313]. These extraction technologies effectively inhibit the thermal decomposition, oxidation, and isomerization of polyphenols while maintaining high extraction yields. Importantly, the structural integrity of polyphenols preserved during the extraction stage directly determines their stability during subsequent processing, which clearly demonstrates the critical carry-over effect of extraction process quality on the stability of the final product.
Overall, these modern approaches fulfill the dual goals of high extraction yield and high bioactivity retention via mild, rapid, and targeted isolation. They clearly outperform conventional methods in efficiency, sustainability, and final product quality, providing a solid foundation for industrial translation.
Current Bottlenecks of Novel Extraction Technologies and Industrial Optimization Directions
Despite the promising potential of novel extraction techniques for phenolic compounds, every single technology still suffers from inherent limitations that hinder its industrial application. The UAE often faces uneven ultrasonic field distribution, leading to inconsistent extraction efficiency across large-scale reactors. MAE tends to generate localized hot spots, which may degrade thermolabile phenolics. SFE involves high equipment and operational costs, limiting its economic viability. NADES, though green, exhibit high viscosity and pose solvent residue challenges that complicate downstream processing. EAE remains constrained by the high cost of commercial enzyme preparations and their limited reusability.
For successful industrial translation, three major aspects of adaptation and optimization are critical. First, equipment development must focus on designing industrial-scale uniform energy field reactors. Second, process standardization is urgently needed. Industrial-scale process parameters (power density, residence time, solvent-to-feed ratio) cannot be directly scaled up from laboratory data; therefore, systematic kinetic modeling and validated amplification protocols are required to establish reliable manufacturing guidelines. Third, cost control remains decisive for economic feasibility. Low-cost, recyclable enzyme preparations and efficient separation-purification technologies for green solvents must be developed to reduce overall production expenses.
Stability Enhancement Strategies for Polyphenol Processing and Application Stages
Polyphenols, as core natural functional active factors in food, face multiple environmental stresses and process disturbances throughout the entire chain of food industrial processing, circulation, and end applications. They are highly prone to oxidative degradation, structural isomerization, covalent binding, and chemical transformation, ultimately resulting in a sharp reduction in content and significant quenching of biological activity. This has become a key scientific bottleneck restricting the high-value utilization of polyphenolic functional factors and the precision development of functional foods.
Core Triggers of Polyphenol Degradation During Processing and Application
The loss of bioactivity and structural integrity of polyphenols during processing, storage, and gastrointestinal transit is primarily attributed to three interdependent degradation drivers: temperature, pH, and oxidation. Understanding how these factors act individually and synergistically is essential for designing stabilization strategies.
Temperature is the primary driver of polyphenol degradation and instability during food processing, exerting destructive effects through two distinct mechanisms. Moderate-to-high temperatures significantly activate endogenous plant and microbial PPO, catalyzing phenol oxidation to quinones that polymerize into inactive condensates. Meanwhile, thermal energy directly cleaves polyphenol glycosidic bonds, releasing less stable free aglycones. Furthermore, high temperature disrupts plant cell substructure integrity, increases polyphenol contact with oxygen and water, and significantly amplifies degradation [100100Alean J, Chejne F, Rojano B. Degradation of polyphenols during cocoa drying. J Food Eng. 2016;189:99-105. Available from: https://doi.org/10.1016/j.jfoodeng.2016.05.026,114114Wang H, Fu Y, Zhao Q. Processing effects on millet polyphenols and anti-diabetic potential. Front Nutr. 2022;9:780499. Available from: https://doi.org/10.3389/fnut.2022.780499]. pH modulates the chemical stability of polyphenols in aqueous systems by reshaping their redox states and existing forms. Alkaline conditions significantly promote the oxidative dehydrogenation of polyphenols to quinones, triggering subsequent molecular condensation. Conversely, acidic conditions inhibit quinone formation, reduce a small fraction of preformed quinones back to parent polyphenols, and enhance stability by maintaining redox equilibrium [115115Usenko OM, Guseynova VP, Sakevich AI. Polyphenol influence on algae under pH changes. Hydrobiol J. 2008;44:37-44. Available from: https://doi.org/10.1615/HydrobJ.v44.i5.40,116116Zeng L, Ma M, Li C. Stability of tea polyphenols at different pH and temperatures. Int J Food Prop. 2017;20:1-18. Available from: https://doi.org/10.1080/10942912.2014.983605]. Oxidation is the core pathway for the structural deterioration of polyphenols. Under multiple stresses, including oxygen, light irradiation, and catalysis by metal ions (Fe3+, Cu2+), polyphenols-rich in polyhydroxy active sites-are highly susceptible to oxidation into ortho-quinone intermediates, which subsequently form dark-colored polymers or undergo covalent interactions with proteins, polysaccharides, and other macromolecules, ultimately resulting in the loss of their biological activities [117117Li C, Arroo RRJ, Shpigelman A. Aminated polyphenols from Strecker degradation in food processing. Food Chem Int. 2025;1:16-23. Available from: https://doi.org/10.1002/fci2,118118Luo Q, Zhang D, Zhou J. Oxidation of tea polyphenols during black tea fermentation. Food Res Int. 2024;196:115016. Available from: https://doi.org/10.1016/j.foodres.2024.115016].
These three drivers do not act independently but form a positive feedback loop that exacerbates polyphenol loss. Alkaline conditions not only directly accelerate oxidation but also lower auto-oxidation activation energy, rendering the system highly sensitive to even trace oxygen exposure. Oxidation is exothermic, generating localized temperature rises that further activate PPO and accelerate all degradation pathways; elevated temperatures, in turn, enhance oxygen and metal ion diffusion, intensifying oxidative chain reactions. This synergistic temperature-oxidation-pH interaction creates a vicious cycle: once initiated, degradation proceeds exponentially rather than linearly, with a combined effect always exceeding the sum of individual effects.
Additionally, light, particularly ultraviolet (UV) radiation, induces photooxidation via excited-state electron transfer to generate free radicals. Mechanical shear during high-speed mixing, homogenization, or pumping disrupts polyphenol-matrix associations and introduces air bubbles, increasing oxygen availability. Non-specific binding of polyphenols to food or biological matrix components alters their bioactivity. Collectively, polyphenol degradation during processing and application is governed by a complex synergistic network centered on temperature, pH, and oxidation. Effective stabilization requires simultaneous control of all three primary drivers and mitigation of secondary factors, including light, shear, and matrix interactions. Failure to address this synergistic feedback loop will inevitably result in substantial phenolic loss, even if individual parameters appear within acceptable ranges.
Strategies for Enhancing Polyphenol Stability in Food Processing Stages
To preserve polyphenol integrity during food processing, a multi-pronged approach targeting degradation pathways is required. The following strategies address the key drivers identified in Section 3.1.
Process optimization: Replacing conventional high-temperature processes with low-temperature alternatives is the first line of defense. Techniques such as low-temperature drying, low-temperature pasteurization, and cold plasma or high-pressure processing minimize thermal activation of PPO and thermal cleavage of glycosidic bonds [119119Almeida FDL, Gomes WF, Cavalcante RS, et al. Cold plasma and high-pressure processing effects on orange juice compounds. Food Res Int. 2017;102:282-290. Available from: https://doi.org/10.1016/j.foodres.2017.09.072]. Additionally, exposure to mechanical shear and light should be minimized. Shortening overall processing time reduces cumulative degradation, as many oxidative pathways follow time-dependent kinetics.
Microenvironment modulation: Controlling the chemical environment around polyphenols is highly effective. Adding organic acids such as citric acid, malic acid, or ascorbic acid maintains a weakly acidic microenvironment (pH 3–4). At this pH range, PPO activity is suppressed, and quinone formation is minimized. Furthermore, incorporating metal ion chelators—phytic acid, Ethylenediaminetetraacetic acid (EDTA), or sodium phytate—sequesters trace Fe2+ and Cu2+, which are potent catalysts of oxidative chain reactions [120120Moon KM, Kwon EB, Lee B. Trends in controlling enzymatic browning in fruits and vegetables. Molecules. 2020;25:2754. Available from: https://doi.org/10.3390/molecules25122754]. This dual approach (low pH + chelation) can significantly slow metal-catalyzed autoxidation.
Antioxidant system formulation: Polyphenols can be co-processed with natural antioxidants to create synergistic protective effects. Vitamin C (ascorbic acid) not only acts as a reducing agent that regenerates oxidized polyphenols but also chelates metal ions. Vitamin E (tocopherols) interrupts lipid peroxidation chains in multiphase systems. Rosemary extract, rich in carnosic acid and rosmarinic acid, provides potent radical-scavenging activity [120120Moon KM, Kwon EB, Lee B. Trends in controlling enzymatic browning in fruits and vegetables. Molecules. 2020;25:2754. Available from: https://doi.org/10.3390/molecules25122754]. When formulated together, these antioxidants delay the consumption of polyphenols, effectively “sacrificing” themselves to preserve the target compounds.
Enzyme activity inhibition: Direct inhibition of endogenous oxidases, especially PPO and peroxidase (POD), is a targeted strategy. Common food-grade inhibitors include reducing agents (e.g., ascorbic acid, cysteine, sodium sulfite), acidulants (already discussed), and physical treatments (e.g., blanching or high-pressure processing). Alternatively, natural inhibitors such as 4-hexylresorcinol can be added at low concentrations [121121Ortiz-Ruiz CV, Berna J, Rodriguez-Lopez JN. Tyrosinase-catalyzed hydroxylation and antibrowning mechanisms. J Agric Food Chem. 2015;63:7032-7040. Available from: https://doi.org/10.1021/acs.jafc.5b02523]. By suppressing enzymatic browning at its source, this approach complements the other strategies to achieve comprehensive polyphenol stabilization.
In summary, a combination of low-temperature processing, acidic chelating microenvironments, antioxidant blends, and enzyme inhibitors offers the most robust protection against polyphenol degradation during food processing.
Targeted Optimization of Polyphenol Stability for Different Application Scenarios
A multi-dimensional synergistic stabilization system is required to prevent polyphenol degradation during food processing, integrating process optimization, microenvironment regulation, antioxidant combination, and targeted enzyme inhibition to comprehensively preserve polyphenol structure and biological activity. Low-temperature alternatives to conventional high-temperature processes serve as the first line of defense. Techniques including freeze-drying, vacuum drying, low-temperature pasteurization, cold plasma, and high-pressure processing minimize thermal activation of PPO and thermal cleavage of glycosidic bonds. Additionally, reducing mechanical shear exposure (via optimized homogenization parameters), light exposure (using amber glass or opaque pipelines), and total processing time mitigates time-dependent cumulative degradation [122122Zawawi NAF, Hazmi NAM, How MS. Inactivation of polyphenol oxidase using thermal and non-thermal methods. Appl Sci. 2022;12:1864. Available from: https://doi.org/10.3390/app12041864,123123Chacha JS, Zhang L, Ofoedu CE. Non-thermal food processing methods: mechanisms and applications. Foods. 2021;10:1430. Available from: https://doi.org/10.3390/foods10061430]. Modulating the chemical microenvironment effectively inhibits oxidative degradation. Organic acids (citric, malic, and ascorbic acid) maintain a weakly acidic pH (3–4), suppressing PPO activity and minimizing quinone formation. Combined with metal chelators (phytic acid, EDTA, sodium phytate) that sequester trace Fe2+ and Cu2+—potent catalysts for oxidative chain reactions—the dual synergy of low pH and chelation significantly retards metal-catalyzed autoxidation [124124Toro-Uribe S, Godoy-Chivatá J, Villamizar-Jaimes AR, Perea-Flores MJ, López-Giraldo LJ. Polyphenol oxidase inhibition in cocoa beans. Antioxidants. 2020;9:458. Available from: https://doi.org/10.3390/antiox9060458,125125Yadav AS. Antioxidant and metal chelating properties of spices and acacia. Food Chem Adv. 2023;2:100257. Available from: https://doi.org/10.1016/j.focha.2023.100257]. Co-processing polyphenols with natural antioxidants generates synergistic protection. Vitamin C regenerates oxidized polyphenols and chelates metals; vitamin E interrupts lipid peroxidation chains in heterogeneous systems; rosemary extract (rich in carnosic acid and rosmarinic acid) provides potent free radical scavenging. These antioxidants delay polyphenol consumption via a "sacrificial" mechanism [108108Wagner A, Dussling S, Scansani S, Bach P, Ludwig M, Steingass CB, et al. Juice production systems and storage effects in red-fleshed apples. Molecules. 2022;27:2459. Available from: https://doi.org/10.3390/molecules27082459,126126Nguyen MM, Karboune S. Antioxidant interactions of essential oils and polyphenols. Antioxidants. 2023;12:486. Available from: https://doi.org/10.3390/antiox12020486,127127Liu XY, Wang WZ, Yao SP. Antioxidant enhancement via hydrogen bonding in rosemary compounds. J Phys Chem B. 2024;128:7627-7638. Available from: https://doi.org/10.1021/acs.jpcb.4c02949]. Furthermore, targeted inhibition of endogenous oxidases (particularly PPO and peroxidase, POD) is a critical complementary strategy. Food-grade reducing agents (ascorbic acid, cysteine, sodium sulfite), physical treatments (blanching, high-pressure processing), or low concentrations of natural inhibitors (4-hexylresorcinol, kojic acid) block enzymatic browning at the source [120120Moon KM, Kwon EB, Lee B. Trends in controlling enzymatic browning in fruits and vegetables. Molecules. 2020;25:2754. Available from: https://doi.org/10.3390/molecules25122754,128128Hamdan N, Lee CH, Wong SL. Prevention of enzymatic browning: natural extracts and genome editing. Molecules. 2022;27:1101. Available from: https://doi.org/10.3390/molecules27031101,129129Marrufo-Hernández NA, Nájera H, González Chávez F. Polyphenol oxidase inactivation using metal–organic frameworks in apple juice. Food Chem. 2024;439:138178. Available from: https://doi.org/10.1016/j.foodchem.2023.138178]. Collectively, the synergistic application of these four strategies provides the most robust technical safeguard against polyphenol degradation during food processing.
Integrated Mechanistic Network for Full-Chain Stability Regulation
Our preceding analyses have systematically demonstrated that polyphenol stability is not governed by any single stage but rather arises from the cumulative effects of biosynthetic structural characteristics, extraction conditions, and processing environments. A comprehensive regulatory network has thus emerged:
Biosynthetic level: Genetic background dictates the inherent biosynthetic potential of polyphenols, developmental stages govern the spatiotemporal expression of this potential, and environmental stresses modulate the magnitude of its realization. These three factors interact synergistically to shape the final content and structural profiles of polyphenols in planta.
Extraction level: Polyphenols with higher degrees of glycosylation and hydroxylation, formed during biosynthesis, exhibit greater stability and lower degradation losses during extraction. Extraction methodologies exert a profound influence on polyphenol degradation kinetics. Furthermore, impurities in the extract (e.g., proteins, polysaccharides, and metal ions) directly impact polyphenol stability during subsequent downstream processing.
Processing level: Polyphenol preparations with lower impurity contents demonstrate superior stability under appropriate storage conditions. Active stabilization strategies implemented during manufacturing can further enhance polyphenol stability throughout the entire processing chain.
Collectively, the synergistic optimization of these three interconnected stages enables substantial improvements in both polyphenol content and stability, representing a transformative approach for industrial-scale production of polyphenol-based functional ingredients.
This paper constructs an analytical framework for the stability and content regulation of plant polyphenols throughout their full life cycle, spanning biosynthesis → extraction → processing → application, breaking the limitation of existing reviews that only focus on individual stages. The shikimate-phenylpropanoid pathway is the core of polyphenol biosynthesis. Biotic and abiotic stresses regulate key rate-limiting enzymes through hormone signaling networks, and mild artificial stress is a green strategy for source enrichment of polyphenols. Polyphenol losses during extraction are mainly caused by oxidation, hydrolysis, and isomerization. Novel coupled enhanced extraction technologies can achieve high yield and high bioactivity retention under mild conditions.
Looking forward, three priority research directions are identified: (1) Fundamental mechanistic studies: multi-omics dissection of tissue-specific stress regulatory networks and quantitative structure-degradability relationships; (2) Technological innovation: development of modular, scalable extraction-processing integrated systems with real-time stability monitoring; (3) Industrial translation: establishment of standardized quality metrics and cost-benefit frameworks for polyphenol fortification in commercial products. Collectively, this full-chain approach will unlock the full industrial potential of plant polyphenols and support evidence-based development of the global functional food and health industry.
Author contributions: Conceptualization, Yuna Li, Guangwei Huang, Roger Ruan, Yanling Cheng.; resources, Guangwei Huang, Roger Ruan, Yanling Cheng.; data curation, Yuna Li.; writing—original draft preparation, Yuna Li.; writing—review and editing, Guangwei Huang, Roger Ruan, Yanling Cheng.; visualization, Guangwei Huang, Roger Ruan, Yanling Cheng.; supervision, Guangwei Huang, Roger Ruan, Yanling Cheng.; project administration, Guangwei Huang, Roger Ruan, Yanling Cheng; funding acquisition, Guangwei Huang, Roger Ruan and Yanling Cheng. All authors have read and agreed to the published version of the manuscript.
Funding: “This research was funded by the Almond Board of California, grant number BIO-22-01-RR and BIO-22-01-YC,” and “The APC was funded by Guangwei Huang”.
Data availability statement: Data sharing not applicable. No new data were created or analyzed in this study.
Acknowledgment: The authors gratefully acknowledge the assistance provided by the Figdraw drawing software.
Abbreviations
The following abbreviations are used in this manuscript.
Abbreviations: The following abbreviations are used in this manuscript
Divyajanani S, Harithpriya K, Ganesan K, Ramkumar KM. Dietary polyphenols remodel DNA methylation patterns of NRF2 in chronic disease. Nutrients. 2023;15:3347. Available from: https://doi.org/10.3390/nu15153347
Qi J, Pan Z, Wang X, Zhang N, He G, Jiang X. Research advances of Zanthoxylum bungeanum polyphenols in inflammatory diseases. Front Immunol. 2024;15:1305886. Available from: https://doi.org/10.3389/fimmu.2024.1305886
Chong Y, Kim BG, Park YJ, Yang Y, Lee SW, Lee Y,et al. Production of four flavonoid C-glucosides in Escherichia coli. J Agric Food Chem. 2023;71:5302-5313. Available from: https://doi.org/10.1021/acs.jafc.3c00297
Vajdi M, Karimi A, Hassanizadeh S, Farhangi MA, Bagherniya M, Askari G, et al. Effect of polyphenols against complications of COVID-19: current evidence and potential efficacy. Pharmacol Rep. 2024;76:307-327. Available from: https://doi.org/10.1007/s43440-024-00585-6
Liu W, Cui X, Zhong Y. Phenolic metabolites as therapeutic in inflammation and neoplasms: molecular pathways explaining their efficacy. Pharmacol Res. 2023;193:106812. Available from: https://doi.org/10.1016/j.phrs.2023.106812
Otręba M, Kośmider L, Stojko J, Rzepecka-Stojko A. Cardioprotective activity of selected polyphenols based on epithelial and aortic cell lines: a review. Molecules. 2020;25:5343. Available from: https://doi.org/10.3390/molecules25225343
Dias MC, Pinto DCGA, Silva AMS. Plant flavonoids: chemical characteristics and biological activity. Molecules. 2021;26:5377. Available from: https://doi.org/10.3390/molecules26175377
Medini F, Ksouri R, Msaada K, Legault J. Phenolic compounds from Limonium densiflorum: antioxidant, anti-inflammatory, anticancer, and anti-influenza activities. Int J Environ Health Res. 2025;35:94-104. Available from: https://doi.org/10.1080/09603123.2024.2342572
Rathod NB, Elabed N, Punia S, Ozogul F, Kim S-K, et al. Recent developments in polyphenol applications on human health: a review. Plants. 2023;12:1217. Available from: https://doi.org/10.3390/plants12061217
Moar K, Yadav S, Pant A, et al. Anti-tumor effects of polyphenols via targeting cancer-driving signaling pathways: a review. Indian J Clin Biochem. 2024;39:470-488. Available from: https://doi.org/10.1007/s12291-024-01222-y
Serreli G, Deiana M. Role of dietary polyphenols in the activity and expression of nitric oxide synthases: a review. Antioxidants. 2023;12:147. Available from: https://doi.org/10.3390/antiox12010147
Kumar K, Debnath P, Singh S, Kumar N. An overview of plant phenolics and their involvement in abiotic stress tolerance. Stresses. 2023;3:570-585. Available from: https://doi.org/10.3390/stresses3030040
Zhou Z, Duan Y, Li Y, Zhang P, Li Q, Yu L, et al. CYP98A monooxygenases: a key enzyme family in plant phenolic compound biosynthesis. Hortic Res. 2025;12:uhaf074. Available from: https://doi.org/10.1093/hr/uhaf074
Chen S, Wang X, Cheng Y, Gao H, Chen X. A review of classification, biosynthesis, biological activities, and applications of flavonoids. Molecules. 2023;28:4982. Available from: https://doi.org/10.3390/molecules28134982
Han S, Cai H, Yu H. UV-C regulation of phenolic biosynthesis in peach fruit during storage. LWT. 2023;190:115573. Available from: https://doi.org/10.1016/j.lwt.2023.115573
Wang S, Xu Y, Wang F. Postharvest changes in phenolic and volatile compounds in grapes. Food Chem. 2025;465:141958. Available from: https://doi.org/10.1016/j.foodchem.2024.141958
Molnar M, Jakovljević Kovač M, Pavić V. Diversity, structure, biosynthesis, and extraction of tannins using deep eutectic solvents. Molecules. 2024;29:2615. Available from: https://doi.org/10.3390/molecules29112615
Sharma P, Dhiman T, Negi RS. Molecular mechanisms of skin photoaging and therapeutic advances using polyphenols. S Afr J Bot. 2024;166:466-482. Available from: https://doi.org/10.1016/j.sajb.2024.01.035
Chrostowski PC, Dietrich AM, Suffet IH. Ozone and oxygen induced oxidative coupling of aqueous phenolics. Water Res. 1983;17:1627-1633. Available from: https://doi.org/10.1016/0043-1354(83)90021-0
Spyroudis S. Hydroxyquinones: synthesis and reactivity. Molecules. 2000;5:1291-1330. Available from: https://doi.org/10.3390/51201291
Alcalde B, Granados M, Saurina J. Exploring the antioxidant features of polyphenols by spectroscopic and electrochemical methods. Antioxidants. 2019;8:523-532. Available from: https://doi.org/10.3390/antiox8110523
Bas TG. Dietary polyphenols (flavonoids) derived from plants for therapeutic health: antioxidant performance, ROS, molecular mechanisms, and bioavailability limitations. Int J Mol Sci. 2026;27:1404. Available from: https://doi.org/10.3390/ijms27031404
Rudrapal M, De Oliveira AM, Singh RP. Dietary polyphenols maintain human health through modulation of gut microbiota. Front Pharmacol. 2026;16:1710088. Available from: https://doi.org/10.3389/fphar.2025.1710088
Stromsnes K, Lagzdina R, Olaso-Gonzalez G, et al. Pharmacological properties of polyphenols: bioavailability, mechanisms of action, and biological effects. Biomedicines. 2021;9:1074. Available from: https://doi.org/10.3390/biomedicines9081074
Wang Y, Chen J, He G, Yin L, Liao Y. Unlocking the potential of flavonoid biosynthesis through integrated metabolic engineering. Front Plant Sci. 2025;16:1597007. Available from: https://doi.org/10.3389/fpls.2025.1597007
Chouhan S, Sharma K, Zha J, Guleria S, Koffas MAG. Recent advances in recombinant biosynthesis of polyphenols. Front Microbiol. 2017;8:2259. Available from: https://doi.org/10.3389/fmicb.2017.02259
Tariq H, Asif S, Andleeb A, Hano C, Abbasi BH. Flavonoid production: plant metabolic engineering and de novo microbial production. Metabolites. 2023;13:124. Available from: https://doi.org/10.3390/metabo13010124
Rosa GP, Barreto MC, Seca AML. Ionic liquids and deep eutectic solvents for polyphenol extraction: opportunities and limitations. Int J Mol Sci. 2026;27:3538. Available from: https://doi.org/10.3390/ijms2708353
Fredsgaard M, Fussy A, Nybo GK. Polyphenols in food and food wastes: extraction, isolation, and health applications. Food Chem Mol Sci. 2026;12:100351. Available from: https://doi.org/10.1016/j.fochms.2025.100351
Palos-Hernández A, González-Paramás AM, Santos-Buelga C. Advances in green extraction of polyphenols from plants and food by-products. Molecules. 2024;30:55. Available from: https://doi.org/10.3390/molecules30010055
Tzin V, Malitsky S, Zvi MMB. Expression of bacterial feedback-insensitive DAHP synthase in Arabidopsis and metabolic bottlenecks. New Phytol. 2012;194:430-439. Available from: https://doi.org/:10.1111/j.1469-8137.2012.04052.x
Yuan J, Zhong S, Long Y. Shikimate kinase plays important roles in anthocyanin synthesis in Petunia. Int J Mol Sci. 2022;23:15964. Available from: https://doi.org/10.3390/ijms232415964
Azizyan R, Abdollahi Mandoulakani B. Gene expression and phenolic compounds in Sonchus arvensis under drought stress. Ind Crops Prod. 2024;209:118030. Available from: https://doi.org/10.1016/j.indcrop.2024.118030
Yao L, Wu X, Jiang X. Subcellular compartmentalization in biosynthesis and engineering of plant natural products. Biotechnol Adv. 2023;69:108258. Available from: https://doi.org/10.1016/j.biotechadv.2023.10825
Jeandet P, Sobarzo-Sánchez E, Silva AS, et al. Biocatalytic and green chemistry methods for resveratrol production. Biotechnol Adv. 2020;39:107461. Available from: https://doi.org/10.1016/j.biotechadv.2019.107461
Liu Q, Li S, Ding W. Aphid-induced tobacco resistance and salicylic acid changes in rhizosphere. Eur J Plant Pathol. 2020;157:465-483. Available from: https://doi.org/10.1007/s10658-020-02005-w
Wang X, Zeng L, Liao Y. Herbivore-induced α-farnesene formation in tea plants. Int J Mol Sci. 2019;20:4151. Available from: https://doi.org/10.3390/ijms20174151
Ji N, Wang J, Li Y. PpWRKY70 involvement in disease resistance via phenylpropanoid pathway in peach. Postharvest Biol Technol. 2021;174:111466. Available from: https://doi.org/10.1016/j.postharvbio.2021.111466
Xie M, Zhang J, Yao T. ANGUSTIFOLIA regulates MYB46 and WRKY33 transcription in Arabidopsis. New Phytol. 2020;228:1627-1639. Available from: https://doi.org/:10.1111/nph.16826
Hodaei M, Rahimmalek M, Arzani A. Water stress effects on flavonoid biosynthesis in chrysanthemum. Ind Crops Prod. 2018;120:295-304. Available from: https://doi.org/10.1016/j.indcrop.2018.04.07
Ding X, Zhu X, Zheng W. BTH treatment delays senescence of postharvest pitaya fruit via antioxidant system and phenylpropanoid pathway. Foods. 2021;10:846. Available from: https://doi.org/10.3390/foods10040846
Mouden S, Bac-Molenaar JA, Kappers IF. Elicitor application in strawberry increases plant resilience without yield loss. Front Plant Sci. 2021;12:695908. Available from: https://doi.org/10.3389/fpls.2021.695908
Manoharan B, Qi SS, Dhandapani V. Gene expression profiling reveals enhanced defense responses in invasive weed vs native congener. Int J Mol Sci. 2019;20:4916. Available from: https://doi.org/10.3390/ijms20194916
Duhan L, Kumar D, Pasrija R. Exogenous salicylic acid mitigates Fusarium stress in Vigna mungo. Plant Cell Rep. 2025;44:2. Available from: https://doi.org/10.1007/s00299-024-03394-6
Guo M, Hou J, Li C. Calcium signaling mediates lignin synthesis in pear exocarp. Plant Physiol Biochem. 2022;190:174-183. Available from: https://doi.org/10.1016/j.plaphy.2022.09.00
Zeng Y, Song H, Xia L. Defensive responses of poplars to fungal pathogens. Front Plant Sci. 2023;14:1107583. Available from: https://doi.org/10.3389/fpls.2023.1107583
Pant SR, Irigoyen S, Liu J. Phenylalanine ammonia lyase promotes antiviral defenses in Brachypodium. mBio. 2021;12:e03518-20. Available from: https://doi.org/10.1128/mBio.03518-20
Carbonaro M, Mattera M. Polyphenoloxidase activity in organic vs conventional fruits. Food Chem. 2001;72:419-424. Available from: https://doi.org/10.1016/S0308-8146(00)00248-X
Merino J, Pedreros A, Fischer S. Weed interference affects polyphenol content in quinoa. Chil J Agric Res. 2019;79:405-414. Available from: https://doi.org/10.4067/S0718-58392019000300405
Tyagi K, Maoz I, Kochanek B. Cytokinin effects on phenylpropanoid pathway in grape. Hortic Res. 2021;8:51-65. Available from: https://doi.org/10.1038/s41438-021-00488-0
Chen Z, Zhang L, Peng M. Selenite enhances Chinese flowering cabbage quality via antioxidant regulation. Food Res Int. 2023;163:112229. Available from: https://doi.org/10.1016/j.foodres.2022.112229
Reshi ZA, Ahmad W, Lukatkin AS. Secondary metabolite biosynthesis pathways and environmental influences. Metabolites. 2023;13:895. Available from: https://doi.org/10.3390/metabo13080895
Yeshi K, Crayn D, Ritmejerytė E. Secondary metabolites under abiotic stress and pharmaceutical applications. Molecules. 2022;27:313. Available from: https://doi.org/10.3390/molecules27010313
Wang Y, Fan K, Wang J. Proteomic analysis of Camellia sinensis under drought stress. J Plant Physiol. 2017;219:91-99. Available from: https://doi.org/10.1016/j.jplph.2017.10.001
Ghotbzadeh Kermani S, Saeidi G, Sabzalian MR. Drought stress influences sesamin and polyphenols in sesame. Food Chem. 2019;289:360-368. Available from: https://doi.org/10.1016/j.foodchem.2019.03.004
Xu N, Liu S, Lu Z. Flavonoid accumulation under salt stress in Ginkgo biloba. Plants. 2020;9:1162. Available from: https://doi.org/10.3390/plants9091162
Fraser DP, Sharma A, Fletcher T. UV-B increases flavonoid quercetin in coriander. Sci Rep. 2017;7:17758. Available from: https://doi.org/10.1038/s41598-017-18073-8
Mosadegh H, Trivellini A, Lucchesini M. UV-B effects on sweet basil physiology. Plants. 2019;8:396. Available from: https://doi.org/10.3390/plants8100396
Chen X, Zhang R, Xin Y. Multi-omics analysis of Betula platyphylla under UV-B stress. Ind Crops Prod. 2024;214:118565. Available from: https://doi.org/10.1016/j.indcrop.2024.118565
Izbiańska K, Arasimowicz-Jelonek M, Deckert J. Phenylpropanoid metabolites improve lupine root tolerance to lead stress. Ecotoxicol Environ Saf. 2014;110:61-67. Available from: https://doi.org/:10.1016/j.ecoenv.2014.08.014
González-Mendoza D, Méndez-Trujillo V, Grimaldo-Juárez O, et al. Changes in photochemical efficiency and epidermal polyphenols in Prosopis species exposed to heavy metals. Open Life Sci. 2017;12:373-378. https://doi.org/10.1515/biol-2017-0043
Ren T, Zheng P, Zhang K, et al. GABA enhances polyphenol accumulation and antioxidant activity in tea under heat stress. Plant Physiol Biochem. 2021;159:363-371. Available from: https://doi.org/10.1016/j.plaphy.2021.01.003
Zhu H, Chen C, Zeng J. MicroRNA528 regulates ROS homeostasis in monocots via copper protein genes. New Phytol. 2020;225:385-399. Available from: https://doi.org/10.1111/nph.16130
Ma X, Chen B, Yang L. GhWRKY55 regulates cotton resistance via lignin biosynthesis and jasmonic acid signaling. Ind Crops Prod. 2024;210:118154. Available from: https://doi.org/10.1016/j.indcrop.2024.118154
Talukder P, Dasgupta M, Hazra A. Impact of invasive weeds on secondary metabolites in okra. Biotechnol Appl Biochem. 2025:e2751. Available from: https://doi.org/10.1002/bab.2751
Yu Y, Wang P, Bai Y. GmFBX176 regulates ABA-mediated drought and salt responses in soybean. Environ Exp Bot. 2020;176:104056. Available from: https://doi.org/10.1016/j.envexpbot.2020.104056
Chen S, Wu F, Li Y. MYB4 and CHS1 regulate flavonoid biosynthesis under salinity stress. Front Plant Sci. 2019;10:178. Available from: https://doi.org/10.3389/fpls.2019.00178
Wang X, Dai WW, Liu C. Physiological responses of purple sweet potato under salinity stress. Genes. 2022;13:1350. Available from: https://doi.org/10.3390/genes13081350
Sahin S, Kısa D, Ataklı SB. Heavy metal effects on antioxidant contents in maize. Rom Agric Res. 2022;39:57-66. Available from: https://doi.org/10.59665/rar3906
Herrmann HA, Dyson BC, Miller MAE. Chloroplast metabolic flux signals cold acclimation in Arabidopsis thaliana. Plant Cell Environ. 2021;44:171-185. Available from: https://doi.org/10.1111/pce.13896
Sridhar A, Ponnuchamy M, Kumar PS. Techniques and modeling of polyphenol extraction: a review. Environ Chem Lett. 2021;19:3409-3443. Available from: https://doi.org/10.1007/s10311-021-01217-8
Dobrinčić A, Repajić M, Garofulić IE. Comparison of extraction methods for olive leaf polyphenols. Processes. 2020;8:1008. Available from: https://doi.org/10.3390/pr8091008
Ding Q, Jiang H, Chen Y. Nitrogen protection in ultrasonic-assisted extraction of rapeseed polyphenols. J Food Process Eng. 2019;42:e13104. Available from: https://doi.org/10.1111/jfpe.13104
Sedraoui S, Badr A, Barba MGM. Optimization of ultrahigh-pressure extraction of phenolics from dates. Food Anal Methods. 2020;13:1556-1569. Available from: https://doi.org/10.1007/s12161-020-01764-w
Tan MJ, Li Y, Zhao SQ. Synergistic ultrasound and pulsed electric field extraction of litchi peel polyphenols. Int J Biol Macromol. 2024;260:129613. Available from: https://doi.org/10.1016/j.ijbiomac.2024.129613
Zhou Z, Shao H, Han X. Enzyme-assisted extraction enhancement of polyphenols from Ulmus pumila. Ind Crops Prod. 2017;97:401-408. Available from: https://doi.org/10.1016/j.indcrop.2016.12.060
Avilés-Betanzos KA, Scampicchio M, Ferrentino G. Supercritical fluid extraction of Capsicum chinense phenolics using RSM. Processes. 2023;11:2055. Available from: https://doi.org/10.3390/pr11072055
Li L, Lv J, Wang X. Green extraction of polyphenols from Elaeagnus angustifolia using natural deep eutectic solvents. Molecules. 2024;29:2412. Available from: https://doi.org/10.3390/molecules29112412
Antony A, Farid M. Effect of temperature on polyphenol extraction. Appl Sci. 2022;12:2107. Available from: https://doi.org/10.3390/app12042107
Park SY, Kang TM, Kim MJ. Enzymatic browning reaction of apple juices prepared using different juicing systems. Biosci Biotechnol Biochem. 2018;82:2000-2006. Available from: https://doi.org/10.1080/09168451.2018.149794
Olszowy-Tomczyk M, Paprotny Ł, Wianowska D. Stability of phenolic acids under extraction conditions. Molecules. 2024;29:5861. Available from: https://doi.org/10.3390/molecules29245861
Wang G, Kumar Y. Mechanisms of initial non-enzymatic oxidation of wine: a mini review. J Food Sci. 2024;89:2530-2545. Available from: https://doi.org/10.1111/1750-3841.17038
Mussio C, Garcia-Perez P, Moret E. Natural chelating agents in food stability: mechanisms and applications. Food Chem. 2025;496:146682. Available from: https://doi.org/10.1016/j.foodchem.2025.14668 .
Wang P, Cheng C, Ma Y. Degradation of polyphenols in aqueous extraction systems under ultrasound. Sep Purif Technol. 2020;247:116967. Available from: https://doi.org/10.1016/j.seppur.2020.116967
Arnold M, Gramza-Michałowska A. Enzymatic browning in apple products: inhibition strategies. Compr Rev Food Sci Food Saf. 2022;21:5038-5076. Available from: https://doi.org/10.1111/1541-4337.13059
Tilley A, McHenry MP, McHenry JA. Role of substrates in polyphenol oxidase-mediated browning. Curr Res Food Sci. 2023;7:100623. Available from: https://doi.org/10.1016/j.crfs.2023.100623
Ebrahimi P, Bayram I, Lante A. Phenolic extract of parsley inhibits lipid oxidation in emulsions. Food Res Int. 2024;187:114452. Available from: doi:10.1016/j.foodres.2024.114452.
Oerlemans K, Barrett DM, Suades CB. Thermal degradation of glucosinolates in red cabbage. Food Chem. 2006;95:19-29. Available from: https://doi.org/10.1016/j.foodchem.2004.12.013
Soong Y, Barlow P. Quantification of gallic and ellagic acid in fruit seeds and kernels. Food Chem. 2006;97:524-530. Available from: https://doi.org/10.1016/j.foodchem.2005.05.033
Ikeda T, Masuda T, Takayama M. Solvent-induced emission of organogels. Org Biomol Chem. 2016;14:36-39. Available from: https://doi.org/10.1039/C5OB01898F
Herrera-Pool E, Ramos-Díaz AL, Lizardi-Jiménez MA. Solvent polarity in ultrasound-assisted extraction of phenolics from habanero pepper leaves. Ultrason Sonochem. 2021;76:105658. Available from: https://doi.org/10.1016/j.ultsonch.2021.105658
Wibisono Y, Rachmawati SA, Mylani VS. Cellulose acetate membrane with Olea europaea nanosolids. Alexandria Eng J. 2023;64:119-129. Available from: https://doi.org/10.1016/j.aej.2022.08.036
Dilokpimol A, Mäkelä MR, Aguilar-Pontes MV. Diversity of fungal feruloyl esterases and applications. Biotechnol Biofuels. 2016;9:231. Available from: https://doi.org/10.1186/s13068-016-0651-6
Gulsunoglu-Konuskan Z, Kilic-Akyilmaz M. Microbial bioconversion of phenolic compounds in agro-industrial wastes. J Agric Food Chem. 2022;70:6901-6910. Available from: https://doi.org/10.1021/acs.jafc.1c06888
Méndez DA, Fabra MJ, Odriozola-Serrano I. Extraction effects on pectin and phenolics from persimmon waste streams. Food Hydrocolloids. 2022;123:107066. Available from: https://doi.org/10.1016/j.foodhyd.2021.107066
Mahamoud R, Bowman DT, Ward WE. Stability of polyphenols in red rooibos tea. Food Chem. 2024;448:139068. Available from: https://doi.org/10.1016/j.foodchem.2024.139068
Fu X, Du Y, Zou L, et al. Acidified glycerol extraction and stabilization of blueberry anthocyanins. Food Chem. 2022;390:133226. Available from: https://doi.org/10.1016/j.foodchem.2022.133226
Hosseini Taheri SE, Bazargan M, Rahnama Vosough P. Composition and oxidation of peanut: a review. J Food Compos Anal. 2024;125:105770. Available from: https://doi.org/10.1016/j.jfca.2023.105770
Alean J, Chejne F, Rojano B. Degradation of polyphenols during cocoa drying. J Food Eng. 2016;189:99-105. Available from: https://doi.org/10.1016/j.jfoodeng.2016.05.026
Leangnim N, Unban K, Thangsunan P. Ultrasonic-assisted enzymatic improvement of polyphenol content and antioxidant activity of miang extracts. Ultrason Sonochem. 2023;94:106351. Available from: https://doi.org/10.1016/j.ultsonch.2023.106351
Chen J, Wang H. Processing characteristics of chia seed: changes in physicochemical properties and structure. Food Biosci. 2024;59:103813. Available from: https://doi.org/10.1016/j.fbio.2024.103813
Kotsou K, Stoikou M, Athanasiadis V. Optimization of extraction to enhance antioxidant properties of Prunus spinosa Horticulturae. 2023;9:942-957.Available from: https://doi.org/10.3390/horticulturae9080942
Chamali S, Bendaoud H, Bouajila J. Optimization of accelerated solvent extraction of bioactive compounds from Eucalyptus intertexta. J Appl Res Med Aromat Plants. 2023;35:100464. Available from: https://doi.org/10.1016/j.jarmap.2023.100464
Wang D, Wang Y, Zhang Z. Degradation and isomerization of dicaffeoylquinic acids under ultrasound. Ultrason Sonochem. 2023;95:106401. Available from: https://doi.org/10.1016/j.ultsonch.2023.106401
Xie C, Yu K, Zhong D, Yuan T, Ye F, Jarrell JA, et al. Isomeric transformations of chlorogenic acid under different conditions. J Agric Food Chem. 2011;59:11078-11087. Available from: https://doi.org/10.1021/jf203104k
Gao Y, Song O, Wang M, Guo X, Zhang G, Liu X, et al. Hydrogen protection enhances bioactivity of chrysanthemum extract. Antioxidants. 2023;12:1019. Available from: https://doi.org/10.3390/antiox12051019
Wagner A, Dussling S, Scansani S, Bach P, Ludwig M, Steingass CB, et al. Juice production systems and storage effects in red-fleshed apples. Molecules. 2022;27:2459. Available from: https://doi.org/10.3390/molecules27082459
Brewer MS. Natural antioxidants: sources, mechanisms, and applications. Compr Rev Food Sci Food Saf. 2011;10:221-247. Available from: https://doi.org/10.1111/j.1541-4337.2011.00156
Losada-Barreiro S, Sezgin-Bayindir Z, Paiva-Martins F, Bravo-Díaz C. Biochemistry of antioxidants: mechanisms and applications. Biomedicines. 2022;10:3051. Available from: https://doi.org/10.3390/biomedicines10123051
Vladić J, Jakovljević Kovač M, Pavić V. Green biomass valorization using supercritical fluids and deep eutectic solvents. Antibiotics. 2023;12:1031. Available from: https://doi.org/10.3390/antibiotics12061031
Lojková L, Pluháčková H, Benešová K. Trends in quercetin extraction methods. TrAC Trends Anal Chem. 2023;167:117229. Available from: https://doi.org/10.1016/j.trac.2023.11722
García-Roldán A, Piriou L, Jauregi P. Natural deep eutectic solvents for polyphenol extraction from coffee grounds. Front Plant Sci. 2023;13:1072592. Available from: https://doi.org/10.3389/fpls.2022.1072592
Wang H, Fu Y, Zhao Q. Processing effects on millet polyphenols and anti-diabetic potential. Front Nutr. 2022;9:780499. Available from: https://doi.org/10.3389/fnut.2022.780499
Usenko OM, Guseynova VP, Sakevich AI. Polyphenol influence on algae under pH changes. Hydrobiol J. 2008;44:37-44. Available from: https://doi.org/10.1615/HydrobJ.v44.i5.40
Zeng L, Ma M, Li C. Stability of tea polyphenols at different pH and temperatures. Int J Food Prop. 2017;20:1-18. Available from: https://doi.org/10.1080/10942912.2014.983605
Li C, Arroo RRJ, Shpigelman A. Aminated polyphenols from Strecker degradation in food processing. Food Chem Int. 2025;1:16-23. Available from: https://doi.org/10.1002/fci2
Luo Q, Zhang D, Zhou J. Oxidation of tea polyphenols during black tea fermentation. Food Res Int. 2024;196:115016. Available from: https://doi.org/10.1016/j.foodres.2024.115016
Almeida FDL, Gomes WF, Cavalcante RS, et al. Cold plasma and high-pressure processing effects on orange juice compounds. Food Res Int. 2017;102:282-290. Available from: https://doi.org/10.1016/j.foodres.2017.09.072
Moon KM, Kwon EB, Lee B. Trends in controlling enzymatic browning in fruits and vegetables. Molecules. 2020;25:2754. Available from: https://doi.org/10.3390/molecules25122754
Ortiz-Ruiz CV, Berna J, Rodriguez-Lopez JN. Tyrosinase-catalyzed hydroxylation and antibrowning mechanisms. J Agric Food Chem. 2015;63:7032-7040. Available from: https://doi.org/10.1021/acs.jafc.5b02523
Zawawi NAF, Hazmi NAM, How MS. Inactivation of polyphenol oxidase using thermal and non-thermal methods. Appl Sci. 2022;12:1864. Available from: https://doi.org/10.3390/app12041864
Chacha JS, Zhang L, Ofoedu CE. Non-thermal food processing methods: mechanisms and applications. Foods. 2021;10:1430. Available from: https://doi.org/10.3390/foods10061430
Yadav AS. Antioxidant and metal chelating properties of spices and acacia. Food Chem Adv. 2023;2:100257. Available from: https://doi.org/10.1016/j.focha.2023.100257
Nguyen MM, Karboune S. Antioxidant interactions of essential oils and polyphenols. Antioxidants. 2023;12:486. Available from: https://doi.org/10.3390/antiox12020486
Liu XY, Wang WZ, Yao SP. Antioxidant enhancement via hydrogen bonding in rosemary compounds. J Phys Chem B. 2024;128:7627-7638. Available from: https://doi.org/10.1021/acs.jpcb.4c02949
Hamdan N, Lee CH, Wong SL. Prevention of enzymatic browning: natural extracts and genome editing. Molecules. 2022;27:1101. Available from: https://doi.org/10.3390/molecules27031101
Marrufo-Hernández NA, Nájera H, González Chávez F. Polyphenol oxidase inactivation using metal–organic frameworks in apple juice. Food Chem. 2024;439:138178. Available from: https://doi.org/10.1016/j.foodchem.2023.138178
Li Y, Huang G, Ruan R, Cheng Y. Stabilization and Valorization of Plant Polyphenols: From Biosynthesis Regulation to Processing and Application. IgMin Res. June 03, 2026; 4(6): 175-190. IgMin ID: igmin343; DOI:10.61927/igmin343; Available at: igmin.link/p343
1College of Biochemical Engineering, Beijing Union University, Beijing, China
2Almond Board of California, Modesto, CA 95354, USA
3Center for Biorefining and Department of Bioproducts and Biosystems Engineering, University of Minnesota, St. Paul, MN 55108, USA
Address Correspondence: Yanling Cheng, College of Biochemical Engineering, Beijing Union University, Beijing, China, Email: [email protected]
How to cite this article: Li Y, Huang G, Ruan R, Cheng Y. Stabilization and Valorization of Plant Polyphenols: From Biosynthesis Regulation to Processing and Application. IgMin Res. June 03, 2026; 4(6): 175-190. IgMin ID: igmin343; DOI:10.61927/igmin343; Available at: igmin.link/p343
Copyright: 2026 Li Y, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Figure 1: Comprehensive biosynthetic pathway network of plan...
Figure 2: Signaling regulatory network of polyphenol biosynt...
Table 1: Types and main changes of biotic stress and abioti...
Table 2: Advanced extraction technology and the yield of po...
Abbreviations: The following abbreviations are used in this manus...
Divyajanani S, Harithpriya K, Ganesan K, Ramkumar KM. Dietary polyphenols remodel DNA methylation patterns of NRF2 in chronic disease. Nutrients. 2023;15:3347. Available from: https://doi.org/10.3390/nu15153347
Qi J, Pan Z, Wang X, Zhang N, He G, Jiang X. Research advances of Zanthoxylum bungeanum polyphenols in inflammatory diseases. Front Immunol. 2024;15:1305886. Available from: https://doi.org/10.3389/fimmu.2024.1305886
Chong Y, Kim BG, Park YJ, Yang Y, Lee SW, Lee Y,et al. Production of four flavonoid C-glucosides in Escherichia coli. J Agric Food Chem. 2023;71:5302-5313. Available from: https://doi.org/10.1021/acs.jafc.3c00297
Vajdi M, Karimi A, Hassanizadeh S, Farhangi MA, Bagherniya M, Askari G, et al. Effect of polyphenols against complications of COVID-19: current evidence and potential efficacy. Pharmacol Rep. 2024;76:307-327. Available from: https://doi.org/10.1007/s43440-024-00585-6
Liu W, Cui X, Zhong Y. Phenolic metabolites as therapeutic in inflammation and neoplasms: molecular pathways explaining their efficacy. Pharmacol Res. 2023;193:106812. Available from: https://doi.org/10.1016/j.phrs.2023.106812
Otręba M, Kośmider L, Stojko J, Rzepecka-Stojko A. Cardioprotective activity of selected polyphenols based on epithelial and aortic cell lines: a review. Molecules. 2020;25:5343. Available from: https://doi.org/10.3390/molecules25225343
Dias MC, Pinto DCGA, Silva AMS. Plant flavonoids: chemical characteristics and biological activity. Molecules. 2021;26:5377. Available from: https://doi.org/10.3390/molecules26175377
Medini F, Ksouri R, Msaada K, Legault J. Phenolic compounds from Limonium densiflorum: antioxidant, anti-inflammatory, anticancer, and anti-influenza activities. Int J Environ Health Res. 2025;35:94-104. Available from: https://doi.org/10.1080/09603123.2024.2342572
Rathod NB, Elabed N, Punia S, Ozogul F, Kim S-K, et al. Recent developments in polyphenol applications on human health: a review. Plants. 2023;12:1217. Available from: https://doi.org/10.3390/plants12061217
Moar K, Yadav S, Pant A, et al. Anti-tumor effects of polyphenols via targeting cancer-driving signaling pathways: a review. Indian J Clin Biochem. 2024;39:470-488. Available from: https://doi.org/10.1007/s12291-024-01222-y
Serreli G, Deiana M. Role of dietary polyphenols in the activity and expression of nitric oxide synthases: a review. Antioxidants. 2023;12:147. Available from: https://doi.org/10.3390/antiox12010147
Kumar K, Debnath P, Singh S, Kumar N. An overview of plant phenolics and their involvement in abiotic stress tolerance. Stresses. 2023;3:570-585. Available from: https://doi.org/10.3390/stresses3030040
Zhou Z, Duan Y, Li Y, Zhang P, Li Q, Yu L, et al. CYP98A monooxygenases: a key enzyme family in plant phenolic compound biosynthesis. Hortic Res. 2025;12:uhaf074. Available from: https://doi.org/10.1093/hr/uhaf074
Chen S, Wang X, Cheng Y, Gao H, Chen X. A review of classification, biosynthesis, biological activities, and applications of flavonoids. Molecules. 2023;28:4982. Available from: https://doi.org/10.3390/molecules28134982
Han S, Cai H, Yu H. UV-C regulation of phenolic biosynthesis in peach fruit during storage. LWT. 2023;190:115573. Available from: https://doi.org/10.1016/j.lwt.2023.115573
Wang S, Xu Y, Wang F. Postharvest changes in phenolic and volatile compounds in grapes. Food Chem. 2025;465:141958. Available from: https://doi.org/10.1016/j.foodchem.2024.141958
Molnar M, Jakovljević Kovač M, Pavić V. Diversity, structure, biosynthesis, and extraction of tannins using deep eutectic solvents. Molecules. 2024;29:2615. Available from: https://doi.org/10.3390/molecules29112615
Sharma P, Dhiman T, Negi RS. Molecular mechanisms of skin photoaging and therapeutic advances using polyphenols. S Afr J Bot. 2024;166:466-482. Available from: https://doi.org/10.1016/j.sajb.2024.01.035
Chrostowski PC, Dietrich AM, Suffet IH. Ozone and oxygen induced oxidative coupling of aqueous phenolics. Water Res. 1983;17:1627-1633. Available from: https://doi.org/10.1016/0043-1354(83)90021-0
Spyroudis S. Hydroxyquinones: synthesis and reactivity. Molecules. 2000;5:1291-1330. Available from: https://doi.org/10.3390/51201291
Alcalde B, Granados M, Saurina J. Exploring the antioxidant features of polyphenols by spectroscopic and electrochemical methods. Antioxidants. 2019;8:523-532. Available from: https://doi.org/10.3390/antiox8110523
Bas TG. Dietary polyphenols (flavonoids) derived from plants for therapeutic health: antioxidant performance, ROS, molecular mechanisms, and bioavailability limitations. Int J Mol Sci. 2026;27:1404. Available from: https://doi.org/10.3390/ijms27031404
Rudrapal M, De Oliveira AM, Singh RP. Dietary polyphenols maintain human health through modulation of gut microbiota. Front Pharmacol. 2026;16:1710088. Available from: https://doi.org/10.3389/fphar.2025.1710088
Stromsnes K, Lagzdina R, Olaso-Gonzalez G, et al. Pharmacological properties of polyphenols: bioavailability, mechanisms of action, and biological effects. Biomedicines. 2021;9:1074. Available from: https://doi.org/10.3390/biomedicines9081074
Wang Y, Chen J, He G, Yin L, Liao Y. Unlocking the potential of flavonoid biosynthesis through integrated metabolic engineering. Front Plant Sci. 2025;16:1597007. Available from: https://doi.org/10.3389/fpls.2025.1597007
Chouhan S, Sharma K, Zha J, Guleria S, Koffas MAG. Recent advances in recombinant biosynthesis of polyphenols. Front Microbiol. 2017;8:2259. Available from: https://doi.org/10.3389/fmicb.2017.02259
Tariq H, Asif S, Andleeb A, Hano C, Abbasi BH. Flavonoid production: plant metabolic engineering and de novo microbial production. Metabolites. 2023;13:124. Available from: https://doi.org/10.3390/metabo13010124
Rosa GP, Barreto MC, Seca AML. Ionic liquids and deep eutectic solvents for polyphenol extraction: opportunities and limitations. Int J Mol Sci. 2026;27:3538. Available from: https://doi.org/10.3390/ijms2708353
Fredsgaard M, Fussy A, Nybo GK. Polyphenols in food and food wastes: extraction, isolation, and health applications. Food Chem Mol Sci. 2026;12:100351. Available from: https://doi.org/10.1016/j.fochms.2025.100351
Palos-Hernández A, González-Paramás AM, Santos-Buelga C. Advances in green extraction of polyphenols from plants and food by-products. Molecules. 2024;30:55. Available from: https://doi.org/10.3390/molecules30010055
Tzin V, Malitsky S, Zvi MMB. Expression of bacterial feedback-insensitive DAHP synthase in Arabidopsis and metabolic bottlenecks. New Phytol. 2012;194:430-439. Available from: https://doi.org/:10.1111/j.1469-8137.2012.04052.x
Yuan J, Zhong S, Long Y. Shikimate kinase plays important roles in anthocyanin synthesis in Petunia. Int J Mol Sci. 2022;23:15964. Available from: https://doi.org/10.3390/ijms232415964
Azizyan R, Abdollahi Mandoulakani B. Gene expression and phenolic compounds in Sonchus arvensis under drought stress. Ind Crops Prod. 2024;209:118030. Available from: https://doi.org/10.1016/j.indcrop.2024.118030
Yao L, Wu X, Jiang X. Subcellular compartmentalization in biosynthesis and engineering of plant natural products. Biotechnol Adv. 2023;69:108258. Available from: https://doi.org/10.1016/j.biotechadv.2023.10825
Jeandet P, Sobarzo-Sánchez E, Silva AS, et al. Biocatalytic and green chemistry methods for resveratrol production. Biotechnol Adv. 2020;39:107461. Available from: https://doi.org/10.1016/j.biotechadv.2019.107461
Liu Q, Li S, Ding W. Aphid-induced tobacco resistance and salicylic acid changes in rhizosphere. Eur J Plant Pathol. 2020;157:465-483. Available from: https://doi.org/10.1007/s10658-020-02005-w
Wang X, Zeng L, Liao Y. Herbivore-induced α-farnesene formation in tea plants. Int J Mol Sci. 2019;20:4151. Available from: https://doi.org/10.3390/ijms20174151
Ji N, Wang J, Li Y. PpWRKY70 involvement in disease resistance via phenylpropanoid pathway in peach. Postharvest Biol Technol. 2021;174:111466. Available from: https://doi.org/10.1016/j.postharvbio.2021.111466
Xie M, Zhang J, Yao T. ANGUSTIFOLIA regulates MYB46 and WRKY33 transcription in Arabidopsis. New Phytol. 2020;228:1627-1639. Available from: https://doi.org/:10.1111/nph.16826
Hodaei M, Rahimmalek M, Arzani A. Water stress effects on flavonoid biosynthesis in chrysanthemum. Ind Crops Prod. 2018;120:295-304. Available from: https://doi.org/10.1016/j.indcrop.2018.04.07
Ding X, Zhu X, Zheng W. BTH treatment delays senescence of postharvest pitaya fruit via antioxidant system and phenylpropanoid pathway. Foods. 2021;10:846. Available from: https://doi.org/10.3390/foods10040846
Mouden S, Bac-Molenaar JA, Kappers IF. Elicitor application in strawberry increases plant resilience without yield loss. Front Plant Sci. 2021;12:695908. Available from: https://doi.org/10.3389/fpls.2021.695908
Manoharan B, Qi SS, Dhandapani V. Gene expression profiling reveals enhanced defense responses in invasive weed vs native congener. Int J Mol Sci. 2019;20:4916. Available from: https://doi.org/10.3390/ijms20194916
Duhan L, Kumar D, Pasrija R. Exogenous salicylic acid mitigates Fusarium stress in Vigna mungo. Plant Cell Rep. 2025;44:2. Available from: https://doi.org/10.1007/s00299-024-03394-6
Guo M, Hou J, Li C. Calcium signaling mediates lignin synthesis in pear exocarp. Plant Physiol Biochem. 2022;190:174-183. Available from: https://doi.org/10.1016/j.plaphy.2022.09.00
Zeng Y, Song H, Xia L. Defensive responses of poplars to fungal pathogens. Front Plant Sci. 2023;14:1107583. Available from: https://doi.org/10.3389/fpls.2023.1107583
Pant SR, Irigoyen S, Liu J. Phenylalanine ammonia lyase promotes antiviral defenses in Brachypodium. mBio. 2021;12:e03518-20. Available from: https://doi.org/10.1128/mBio.03518-20
Carbonaro M, Mattera M. Polyphenoloxidase activity in organic vs conventional fruits. Food Chem. 2001;72:419-424. Available from: https://doi.org/10.1016/S0308-8146(00)00248-X
Merino J, Pedreros A, Fischer S. Weed interference affects polyphenol content in quinoa. Chil J Agric Res. 2019;79:405-414. Available from: https://doi.org/10.4067/S0718-58392019000300405
Tyagi K, Maoz I, Kochanek B. Cytokinin effects on phenylpropanoid pathway in grape. Hortic Res. 2021;8:51-65. Available from: https://doi.org/10.1038/s41438-021-00488-0
Chen Z, Zhang L, Peng M. Selenite enhances Chinese flowering cabbage quality via antioxidant regulation. Food Res Int. 2023;163:112229. Available from: https://doi.org/10.1016/j.foodres.2022.112229
Reshi ZA, Ahmad W, Lukatkin AS. Secondary metabolite biosynthesis pathways and environmental influences. Metabolites. 2023;13:895. Available from: https://doi.org/10.3390/metabo13080895
Yeshi K, Crayn D, Ritmejerytė E. Secondary metabolites under abiotic stress and pharmaceutical applications. Molecules. 2022;27:313. Available from: https://doi.org/10.3390/molecules27010313
Wang Y, Fan K, Wang J. Proteomic analysis of Camellia sinensis under drought stress. J Plant Physiol. 2017;219:91-99. Available from: https://doi.org/10.1016/j.jplph.2017.10.001
Ghotbzadeh Kermani S, Saeidi G, Sabzalian MR. Drought stress influences sesamin and polyphenols in sesame. Food Chem. 2019;289:360-368. Available from: https://doi.org/10.1016/j.foodchem.2019.03.004
Xu N, Liu S, Lu Z. Flavonoid accumulation under salt stress in Ginkgo biloba. Plants. 2020;9:1162. Available from: https://doi.org/10.3390/plants9091162
Fraser DP, Sharma A, Fletcher T. UV-B increases flavonoid quercetin in coriander. Sci Rep. 2017;7:17758. Available from: https://doi.org/10.1038/s41598-017-18073-8
Mosadegh H, Trivellini A, Lucchesini M. UV-B effects on sweet basil physiology. Plants. 2019;8:396. Available from: https://doi.org/10.3390/plants8100396
Chen X, Zhang R, Xin Y. Multi-omics analysis of Betula platyphylla under UV-B stress. Ind Crops Prod. 2024;214:118565. Available from: https://doi.org/10.1016/j.indcrop.2024.118565
Izbiańska K, Arasimowicz-Jelonek M, Deckert J. Phenylpropanoid metabolites improve lupine root tolerance to lead stress. Ecotoxicol Environ Saf. 2014;110:61-67. Available from: https://doi.org/:10.1016/j.ecoenv.2014.08.014
González-Mendoza D, Méndez-Trujillo V, Grimaldo-Juárez O, et al. Changes in photochemical efficiency and epidermal polyphenols in Prosopis species exposed to heavy metals. Open Life Sci. 2017;12:373-378. https://doi.org/10.1515/biol-2017-0043
Ren T, Zheng P, Zhang K, et al. GABA enhances polyphenol accumulation and antioxidant activity in tea under heat stress. Plant Physiol Biochem. 2021;159:363-371. Available from: https://doi.org/10.1016/j.plaphy.2021.01.003
Zhu H, Chen C, Zeng J. MicroRNA528 regulates ROS homeostasis in monocots via copper protein genes. New Phytol. 2020;225:385-399. Available from: https://doi.org/10.1111/nph.16130
Ma X, Chen B, Yang L. GhWRKY55 regulates cotton resistance via lignin biosynthesis and jasmonic acid signaling. Ind Crops Prod. 2024;210:118154. Available from: https://doi.org/10.1016/j.indcrop.2024.118154
Talukder P, Dasgupta M, Hazra A. Impact of invasive weeds on secondary metabolites in okra. Biotechnol Appl Biochem. 2025:e2751. Available from: https://doi.org/10.1002/bab.2751
Yu Y, Wang P, Bai Y. GmFBX176 regulates ABA-mediated drought and salt responses in soybean. Environ Exp Bot. 2020;176:104056. Available from: https://doi.org/10.1016/j.envexpbot.2020.104056
Chen S, Wu F, Li Y. MYB4 and CHS1 regulate flavonoid biosynthesis under salinity stress. Front Plant Sci. 2019;10:178. Available from: https://doi.org/10.3389/fpls.2019.00178
Wang X, Dai WW, Liu C. Physiological responses of purple sweet potato under salinity stress. Genes. 2022;13:1350. Available from: https://doi.org/10.3390/genes13081350
Sahin S, Kısa D, Ataklı SB. Heavy metal effects on antioxidant contents in maize. Rom Agric Res. 2022;39:57-66. Available from: https://doi.org/10.59665/rar3906
Herrmann HA, Dyson BC, Miller MAE. Chloroplast metabolic flux signals cold acclimation in Arabidopsis thaliana. Plant Cell Environ. 2021;44:171-185. Available from: https://doi.org/10.1111/pce.13896
Sridhar A, Ponnuchamy M, Kumar PS. Techniques and modeling of polyphenol extraction: a review. Environ Chem Lett. 2021;19:3409-3443. Available from: https://doi.org/10.1007/s10311-021-01217-8
Dobrinčić A, Repajić M, Garofulić IE. Comparison of extraction methods for olive leaf polyphenols. Processes. 2020;8:1008. Available from: https://doi.org/10.3390/pr8091008
Ding Q, Jiang H, Chen Y. Nitrogen protection in ultrasonic-assisted extraction of rapeseed polyphenols. J Food Process Eng. 2019;42:e13104. Available from: https://doi.org/10.1111/jfpe.13104
Sedraoui S, Badr A, Barba MGM. Optimization of ultrahigh-pressure extraction of phenolics from dates. Food Anal Methods. 2020;13:1556-1569. Available from: https://doi.org/10.1007/s12161-020-01764-w
Tan MJ, Li Y, Zhao SQ. Synergistic ultrasound and pulsed electric field extraction of litchi peel polyphenols. Int J Biol Macromol. 2024;260:129613. Available from: https://doi.org/10.1016/j.ijbiomac.2024.129613
Zhou Z, Shao H, Han X. Enzyme-assisted extraction enhancement of polyphenols from Ulmus pumila. Ind Crops Prod. 2017;97:401-408. Available from: https://doi.org/10.1016/j.indcrop.2016.12.060
Avilés-Betanzos KA, Scampicchio M, Ferrentino G. Supercritical fluid extraction of Capsicum chinense phenolics using RSM. Processes. 2023;11:2055. Available from: https://doi.org/10.3390/pr11072055
Li L, Lv J, Wang X. Green extraction of polyphenols from Elaeagnus angustifolia using natural deep eutectic solvents. Molecules. 2024;29:2412. Available from: https://doi.org/10.3390/molecules29112412
Antony A, Farid M. Effect of temperature on polyphenol extraction. Appl Sci. 2022;12:2107. Available from: https://doi.org/10.3390/app12042107
Park SY, Kang TM, Kim MJ. Enzymatic browning reaction of apple juices prepared using different juicing systems. Biosci Biotechnol Biochem. 2018;82:2000-2006. Available from: https://doi.org/10.1080/09168451.2018.149794
Olszowy-Tomczyk M, Paprotny Ł, Wianowska D. Stability of phenolic acids under extraction conditions. Molecules. 2024;29:5861. Available from: https://doi.org/10.3390/molecules29245861
Wang G, Kumar Y. Mechanisms of initial non-enzymatic oxidation of wine: a mini review. J Food Sci. 2024;89:2530-2545. Available from: https://doi.org/10.1111/1750-3841.17038
Mussio C, Garcia-Perez P, Moret E. Natural chelating agents in food stability: mechanisms and applications. Food Chem. 2025;496:146682. Available from: https://doi.org/10.1016/j.foodchem.2025.14668 .
Wang P, Cheng C, Ma Y. Degradation of polyphenols in aqueous extraction systems under ultrasound. Sep Purif Technol. 2020;247:116967. Available from: https://doi.org/10.1016/j.seppur.2020.116967
Arnold M, Gramza-Michałowska A. Enzymatic browning in apple products: inhibition strategies. Compr Rev Food Sci Food Saf. 2022;21:5038-5076. Available from: https://doi.org/10.1111/1541-4337.13059
Tilley A, McHenry MP, McHenry JA. Role of substrates in polyphenol oxidase-mediated browning. Curr Res Food Sci. 2023;7:100623. Available from: https://doi.org/10.1016/j.crfs.2023.100623
Ebrahimi P, Bayram I, Lante A. Phenolic extract of parsley inhibits lipid oxidation in emulsions. Food Res Int. 2024;187:114452. Available from: doi:10.1016/j.foodres.2024.114452.
Oerlemans K, Barrett DM, Suades CB. Thermal degradation of glucosinolates in red cabbage. Food Chem. 2006;95:19-29. Available from: https://doi.org/10.1016/j.foodchem.2004.12.013
Soong Y, Barlow P. Quantification of gallic and ellagic acid in fruit seeds and kernels. Food Chem. 2006;97:524-530. Available from: https://doi.org/10.1016/j.foodchem.2005.05.033
Ikeda T, Masuda T, Takayama M. Solvent-induced emission of organogels. Org Biomol Chem. 2016;14:36-39. Available from: https://doi.org/10.1039/C5OB01898F
Herrera-Pool E, Ramos-Díaz AL, Lizardi-Jiménez MA. Solvent polarity in ultrasound-assisted extraction of phenolics from habanero pepper leaves. Ultrason Sonochem. 2021;76:105658. Available from: https://doi.org/10.1016/j.ultsonch.2021.105658
Wibisono Y, Rachmawati SA, Mylani VS. Cellulose acetate membrane with Olea europaea nanosolids. Alexandria Eng J. 2023;64:119-129. Available from: https://doi.org/10.1016/j.aej.2022.08.036
Dilokpimol A, Mäkelä MR, Aguilar-Pontes MV. Diversity of fungal feruloyl esterases and applications. Biotechnol Biofuels. 2016;9:231. Available from: https://doi.org/10.1186/s13068-016-0651-6
Gulsunoglu-Konuskan Z, Kilic-Akyilmaz M. Microbial bioconversion of phenolic compounds in agro-industrial wastes. J Agric Food Chem. 2022;70:6901-6910. Available from: https://doi.org/10.1021/acs.jafc.1c06888
Méndez DA, Fabra MJ, Odriozola-Serrano I. Extraction effects on pectin and phenolics from persimmon waste streams. Food Hydrocolloids. 2022;123:107066. Available from: https://doi.org/10.1016/j.foodhyd.2021.107066
Mahamoud R, Bowman DT, Ward WE. Stability of polyphenols in red rooibos tea. Food Chem. 2024;448:139068. Available from: https://doi.org/10.1016/j.foodchem.2024.139068
Fu X, Du Y, Zou L, et al. Acidified glycerol extraction and stabilization of blueberry anthocyanins. Food Chem. 2022;390:133226. Available from: https://doi.org/10.1016/j.foodchem.2022.133226
Hosseini Taheri SE, Bazargan M, Rahnama Vosough P. Composition and oxidation of peanut: a review. J Food Compos Anal. 2024;125:105770. Available from: https://doi.org/10.1016/j.jfca.2023.105770
Alean J, Chejne F, Rojano B. Degradation of polyphenols during cocoa drying. J Food Eng. 2016;189:99-105. Available from: https://doi.org/10.1016/j.jfoodeng.2016.05.026
Leangnim N, Unban K, Thangsunan P. Ultrasonic-assisted enzymatic improvement of polyphenol content and antioxidant activity of miang extracts. Ultrason Sonochem. 2023;94:106351. Available from: https://doi.org/10.1016/j.ultsonch.2023.106351
Chen J, Wang H. Processing characteristics of chia seed: changes in physicochemical properties and structure. Food Biosci. 2024;59:103813. Available from: https://doi.org/10.1016/j.fbio.2024.103813
Kotsou K, Stoikou M, Athanasiadis V. Optimization of extraction to enhance antioxidant properties of Prunus spinosa Horticulturae. 2023;9:942-957.Available from: https://doi.org/10.3390/horticulturae9080942
Chamali S, Bendaoud H, Bouajila J. Optimization of accelerated solvent extraction of bioactive compounds from Eucalyptus intertexta. J Appl Res Med Aromat Plants. 2023;35:100464. Available from: https://doi.org/10.1016/j.jarmap.2023.100464
Wang D, Wang Y, Zhang Z. Degradation and isomerization of dicaffeoylquinic acids under ultrasound. Ultrason Sonochem. 2023;95:106401. Available from: https://doi.org/10.1016/j.ultsonch.2023.106401
Xie C, Yu K, Zhong D, Yuan T, Ye F, Jarrell JA, et al. Isomeric transformations of chlorogenic acid under different conditions. J Agric Food Chem. 2011;59:11078-11087. Available from: https://doi.org/10.1021/jf203104k
Gao Y, Song O, Wang M, Guo X, Zhang G, Liu X, et al. Hydrogen protection enhances bioactivity of chrysanthemum extract. Antioxidants. 2023;12:1019. Available from: https://doi.org/10.3390/antiox12051019
Wagner A, Dussling S, Scansani S, Bach P, Ludwig M, Steingass CB, et al. Juice production systems and storage effects in red-fleshed apples. Molecules. 2022;27:2459. Available from: https://doi.org/10.3390/molecules27082459
Brewer MS. Natural antioxidants: sources, mechanisms, and applications. Compr Rev Food Sci Food Saf. 2011;10:221-247. Available from: https://doi.org/10.1111/j.1541-4337.2011.00156
Losada-Barreiro S, Sezgin-Bayindir Z, Paiva-Martins F, Bravo-Díaz C. Biochemistry of antioxidants: mechanisms and applications. Biomedicines. 2022;10:3051. Available from: https://doi.org/10.3390/biomedicines10123051
Vladić J, Jakovljević Kovač M, Pavić V. Green biomass valorization using supercritical fluids and deep eutectic solvents. Antibiotics. 2023;12:1031. Available from: https://doi.org/10.3390/antibiotics12061031
Lojková L, Pluháčková H, Benešová K. Trends in quercetin extraction methods. TrAC Trends Anal Chem. 2023;167:117229. Available from: https://doi.org/10.1016/j.trac.2023.11722
García-Roldán A, Piriou L, Jauregi P. Natural deep eutectic solvents for polyphenol extraction from coffee grounds. Front Plant Sci. 2023;13:1072592. Available from: https://doi.org/10.3389/fpls.2022.1072592
Wang H, Fu Y, Zhao Q. Processing effects on millet polyphenols and anti-diabetic potential. Front Nutr. 2022;9:780499. Available from: https://doi.org/10.3389/fnut.2022.780499
Usenko OM, Guseynova VP, Sakevich AI. Polyphenol influence on algae under pH changes. Hydrobiol J. 2008;44:37-44. Available from: https://doi.org/10.1615/HydrobJ.v44.i5.40
Zeng L, Ma M, Li C. Stability of tea polyphenols at different pH and temperatures. Int J Food Prop. 2017;20:1-18. Available from: https://doi.org/10.1080/10942912.2014.983605
Li C, Arroo RRJ, Shpigelman A. Aminated polyphenols from Strecker degradation in food processing. Food Chem Int. 2025;1:16-23. Available from: https://doi.org/10.1002/fci2
Luo Q, Zhang D, Zhou J. Oxidation of tea polyphenols during black tea fermentation. Food Res Int. 2024;196:115016. Available from: https://doi.org/10.1016/j.foodres.2024.115016
Almeida FDL, Gomes WF, Cavalcante RS, et al. Cold plasma and high-pressure processing effects on orange juice compounds. Food Res Int. 2017;102:282-290. Available from: https://doi.org/10.1016/j.foodres.2017.09.072
Moon KM, Kwon EB, Lee B. Trends in controlling enzymatic browning in fruits and vegetables. Molecules. 2020;25:2754. Available from: https://doi.org/10.3390/molecules25122754
Ortiz-Ruiz CV, Berna J, Rodriguez-Lopez JN. Tyrosinase-catalyzed hydroxylation and antibrowning mechanisms. J Agric Food Chem. 2015;63:7032-7040. Available from: https://doi.org/10.1021/acs.jafc.5b02523
Zawawi NAF, Hazmi NAM, How MS. Inactivation of polyphenol oxidase using thermal and non-thermal methods. Appl Sci. 2022;12:1864. Available from: https://doi.org/10.3390/app12041864
Chacha JS, Zhang L, Ofoedu CE. Non-thermal food processing methods: mechanisms and applications. Foods. 2021;10:1430. Available from: https://doi.org/10.3390/foods10061430
Yadav AS. Antioxidant and metal chelating properties of spices and acacia. Food Chem Adv. 2023;2:100257. Available from: https://doi.org/10.1016/j.focha.2023.100257
Nguyen MM, Karboune S. Antioxidant interactions of essential oils and polyphenols. Antioxidants. 2023;12:486. Available from: https://doi.org/10.3390/antiox12020486
Liu XY, Wang WZ, Yao SP. Antioxidant enhancement via hydrogen bonding in rosemary compounds. J Phys Chem B. 2024;128:7627-7638. Available from: https://doi.org/10.1021/acs.jpcb.4c02949
Hamdan N, Lee CH, Wong SL. Prevention of enzymatic browning: natural extracts and genome editing. Molecules. 2022;27:1101. Available from: https://doi.org/10.3390/molecules27031101
Marrufo-Hernández NA, Nájera H, González Chávez F. Polyphenol oxidase inactivation using metal–organic frameworks in apple juice. Food Chem. 2024;439:138178. Available from: https://doi.org/10.1016/j.foodchem.2023.138178



 スキャンしてリンクを取得
スキャンしてリンクを取得